 Back
BackATP Synthase, Mitochondrial Transport, and Regulation of Oxidative Phosphorylation
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
ATP Synthase Complex
Structure and Function of ATP Synthase
The ATP synthase complex is a large, multi-subunit enzyme embedded in the inner mitochondrial membrane, responsible for synthesizing ATP from ADP and inorganic phosphate (Pi) using the energy of the proton-motive force generated by the electron transport chain. It is composed of two main components: F0, which forms the proton channel, and F1, which contains the catalytic sites for ATP synthesis.
F0 component: Embedded in the membrane, forms a channel for protons (H+) to flow from the intermembrane space into the mitochondrial matrix.
F1 component: Projects into the matrix and contains the catalytic sites for ATP synthesis, primarily in the β subunits.
Three functional parts: (1) Rotor (c ring and γ subunit), (2) Catalytic headpiece (α/β subunits), (3) Stator (a, b, d, h subunits, and OSCP).
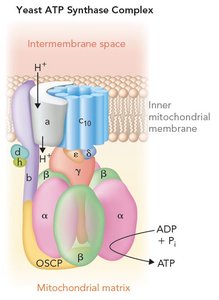
Key Point: Proton flow through F0 drives rotation of the γ subunit, inducing conformational changes in the β subunits of F1 that catalyze ATP synthesis.
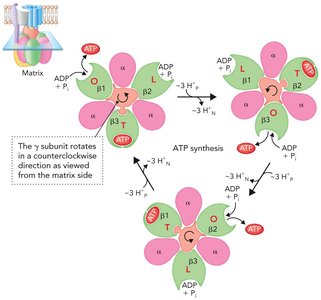
Binding Change Mechanism Model
The binding change mechanism explains how ATP synthase couples proton flow to ATP synthesis. The γ subunit rotates within the α3β3 hexamer, causing each β subunit to cycle through three conformations:
Loose (L): Binds ADP and Pi.
Tight (T): Catalyzes ATP synthesis from ADP and Pi.
Open (O): Releases ATP.
Each 120° rotation of the γ subunit changes the conformation of the β subunits, resulting in the synthesis and release of one ATP per β subunit per full rotation.
Equation:
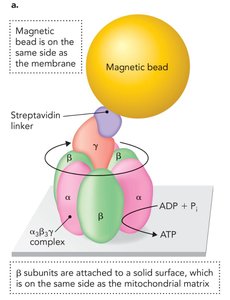
ATP Synthase as a Molecular Motor
ATP synthase can function in reverse as an ATP hydrolase, using ATP hydrolysis to pump protons against their gradient. Experimental setups have demonstrated that forced rotation of the γ subunit (e.g., using magnetic beads or actin filaments) can drive ATP synthesis even in the absence of a proton gradient, highlighting its function as a reversible molecular motor.
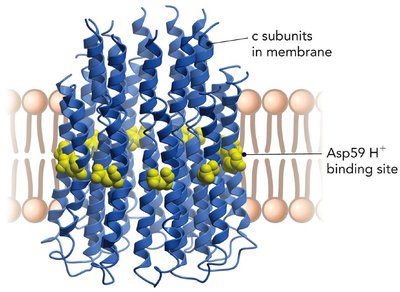
Proton Translocation and γ Subunit Rotation
Proton movement through the c ring of F0 is coupled to the rotation of the γ subunit. The two-channel model proposes that protons bind to a conserved Asp residue in each c subunit, causing conformational changes that drive rotation. This rotation is transmitted to the catalytic headpiece, enabling ATP synthesis.
Mitochondrial Transport Systems
ATP/ADP and Phosphate Translocases
ATP produced in the mitochondrial matrix must be exported to the cytosol, while ADP and Pi must be imported for continued ATP synthesis. This is achieved by two key transport proteins:
ATP/ADP translocase: Exchanges ATP (out) for ADP (in) across the inner mitochondrial membrane. It operates via two conformational states (C and M), which can be inhibited by atractyloside and bongkrekic acid, respectively.
Phosphate translocase: Imports Pi into the matrix, functioning either as a symporter (with H+) or as an antiporter (with OH−).

Table: Inhibitors of ATP/ADP Translocase
Inhibitor | Target State | Source |
|---|---|---|
Atractyloside | C state | Atractylis gummifera (thistle plant) |
Bongkrekic acid | M state | Bacterial contamination in tempeh |
Shuttle Systems for NADH
NADH generated in the cytosol (e.g., during glycolysis) cannot cross the inner mitochondrial membrane. Instead, electrons are transferred into the matrix via shuttle systems:
Malate-Aspartate Shuttle: Used in liver cells; transfers electrons from cytosolic NADH to mitochondrial NAD+ via malate and aspartate intermediates.
Glycerol-3-Phosphate Shuttle: Used in muscle cells; transfers electrons from NADH to FAD, resulting in a lower ATP yield but faster response to energy demands.
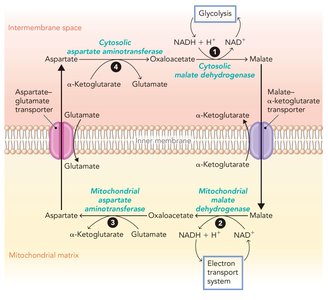
Key Point: The choice of shuttle system affects the net ATP yield from glucose oxidation (32 ATP in liver, 30 ATP in muscle).
Regulation of Oxidative Phosphorylation
Allosteric Regulation
Oxidative phosphorylation is tightly regulated to match cellular energy demands. High levels of ATP and NADH signal high energy charge and act as allosteric inhibitors, decreasing the flux through the electron transport chain and ATP synthase. Conversely, ADP, AMP, and Pi are allosteric activators, increasing ATP production when energy is needed.
ATP and NADH: Inhibit oxidative phosphorylation.
ADP, AMP, Pi: Activate oxidative phosphorylation.
Inhibitors of Oxidative Phosphorylation
Three main classes of inhibitors can decrease mitochondrial ATP synthesis:
Electron transport system inhibitors: Block electron flow (e.g., cyanide).
Uncouplers: Allow protons to cross the membrane without ATP synthesis (e.g., 2,4-dinitrophenol, DNP).
ATP synthase inhibitors: Directly inhibit the ATP synthase complex (e.g., oligomycin).
Example: DNP dissipates the proton gradient, causing increased fat oxidation and heat production but preventing ATP synthesis—a dangerous effect exploited in weight loss but with severe risks.
Inherited Mitochondrial Diseases
Mutations in genes encoding components of the electron transport chain or ATP synthase can lead to mitochondrial diseases, often affecting tissues with high energy demands such as neurons and muscles. These diseases are maternally inherited due to the maternal origin of mitochondria in fertilization.
Commonly affected complexes: Complexes I, III, IV, and the A6 subunit of ATP synthase.
Genetic transmission: Only maternal mitochondria are inherited by offspring.
Summary Table: ATP Yield from Glucose Oxidation
Shuttle System | Cell Type | Net ATP Yield |
|---|---|---|
Malate-Aspartate | Liver | 32 ATP |
Glycerol-3-Phosphate | Muscle | 30 ATP |
Additional info: The ATP synthase complex is a prime example of how protein structure and conformational changes are harnessed to perform essential cellular work, linking the chemical energy of redox reactions to the synthesis of ATP, the universal energy currency of the cell.
