 Back
BackEnzyme Catalysis and Carbohydrate Structure: Core Concepts in Biochemistry
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
Enzyme Catalysis
Definition and General Properties of Catalysts
Catalysts are substances that increase the rate of a chemical reaction without themselves undergoing permanent chemical change. In biological systems, enzymes serve as highly efficient and specific catalysts for biochemical reactions.
Lower Activation Energy: Catalysts lower the activation energy (Ea) required for a reaction, increasing the fraction of molecules that can reach the transition state.
Equilibrium Position: Catalysts do not affect the position of equilibrium; they only accelerate the rate at which equilibrium is achieved.
Bidirectional Effect: Catalysts accelerate both the forward and reverse reactions equally.
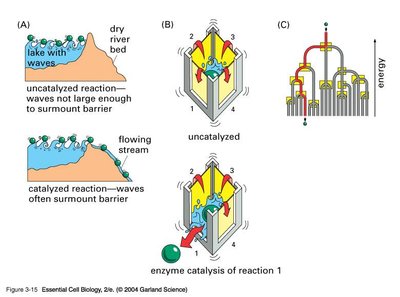
Enzyme Catalysis Mechanisms
Enzymes catalyze reactions by providing an alternative reaction pathway and stabilizing the transition state, thereby reducing the energy required to reach the highest energy state of the reaction.
Transition State Stabilization: Enzymes bind substrates in a way that stabilizes the transition state, lowering the activation energy.
Entropic and Enthalpic Factors: Enzymes enhance reaction rates by positioning and orienting substrates, reducing entropy, and providing favorable enthalpic interactions.
Induced Fit Model: The enzyme undergoes conformational changes upon substrate binding, increasing affinity for the transition state and further stabilizing it.
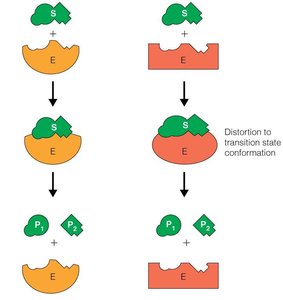
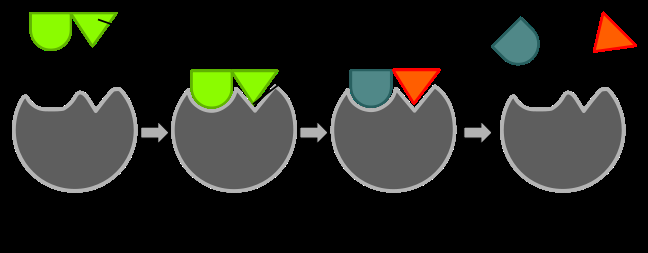
Michaelis-Menten Kinetics
The Michaelis-Menten equation describes the rate of enzymatic reactions as a function of substrate concentration:
Vmax: Maximum reaction velocity at saturating substrate concentration.
Km: Substrate concentration at which the reaction rate is half-maximal; reflects enzyme affinity for substrate.
Kcat: Turnover number, the maximum number of substrate molecules converted to product per enzyme molecule per second.
Catalytic Efficiency: Given by , representing enzyme efficiency at low substrate concentrations.
Amino Acids in Enzyme Active Sites
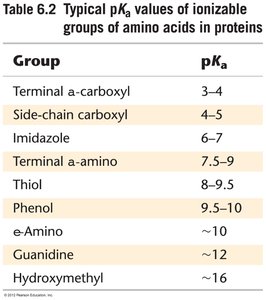
Certain amino acids are frequently found in enzyme active sites due to their ability to participate in catalysis through acid-base reactions, nucleophilic attack, or covalent catalysis. Their pKa values can be significantly shifted in the enzyme environment.
Group | pKa |
|---|---|
Terminal α-carboxyl | 3–4 |
Side-chain carboxyl | 4–5 |
Imidazole | 6–7 |
Terminal α-amino | 7.5–9 |
Thiol | 8.9–9.5 |
Phenol | 9.5–10 |
ε-Amino | ~10 |
Guanidine | ~12 |
Hydroxymethyl | ~16 |
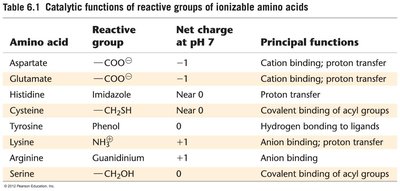
Amino acid | Reactive group | Net charge at pH 7 | Principal functions |
|---|---|---|---|
Aspartate | —COO− | −1 | Cation binding; proton transfer |
Glutamate | —COO− | −1 | Cation binding; proton transfer |
Histidine | Imidazole | Near 0 | Proton transfer |
Cysteine | —CH2SH | Near 0 | Covalent binding of acyl groups |
Tyrosine | Phenol | 0 | Hydrogen bonding to ligands |
Lysine | NH3+ | +1 | Anion binding; proton transfer |
Arginine | Guanidinium | +1 | Anion binding |
Serine | —CH2OH | 0 | Covalent binding of acyl groups |
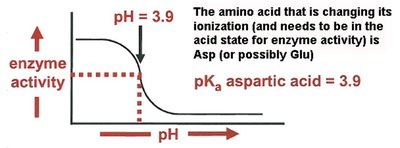
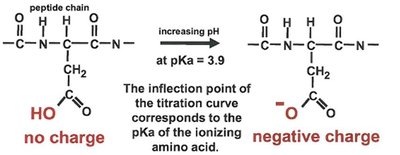
pH Dependence of Enzyme Activity
The activity of enzymes is highly dependent on pH, as changes in pH can alter the ionization state of amino acid side chains critical for catalysis. The pKa of these groups can be determined by titration experiments.
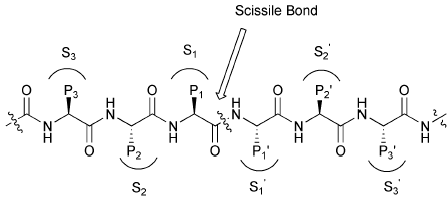
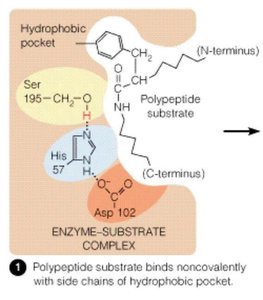
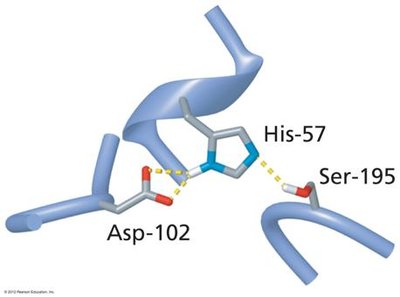
Serine Protease Mechanism and the Catalytic Triad
Serine proteases, such as chymotrypsin, utilize a catalytic triad (Ser195, His57, Asp102) to cleave peptide bonds. The triad works together to activate the serine residue for nucleophilic attack on the peptide bond.
Ser195: Acts as a nucleophile, attacking the carbonyl carbon of the peptide bond.
His57: Functions as a general base, accepting a proton from Ser195.
Asp102: Stabilizes the positively charged His57, enhancing its basicity.
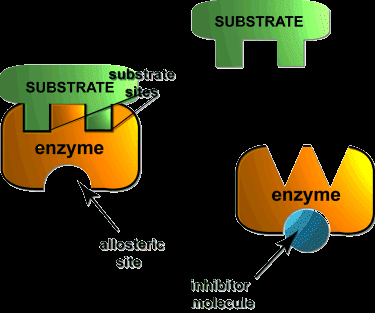
Enzyme Inhibition
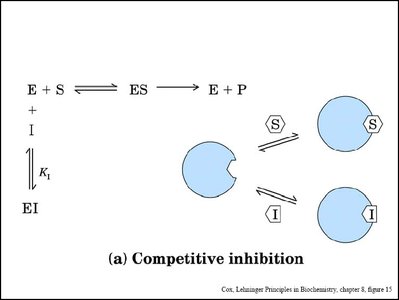
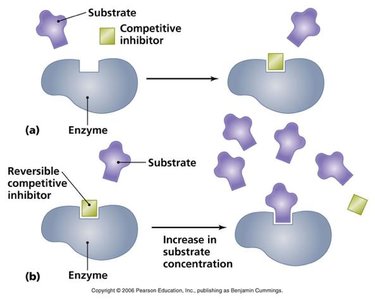
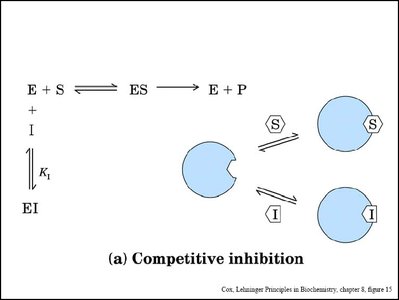
Types of Reversible Inhibition
Competitive Inhibition: Inhibitor binds to the active site, competing with the substrate. Increases apparent Km, Vmax unchanged.
Noncompetitive (Mixed) Inhibition: Inhibitor binds to enzyme or enzyme-substrate complex at a site other than the active site. Decreases Vmax, Km may change.
Uncompetitive Inhibition: Inhibitor binds only to the enzyme-substrate complex, decreasing both Vmax and Km by the same factor.
Regulation of Enzyme Activity
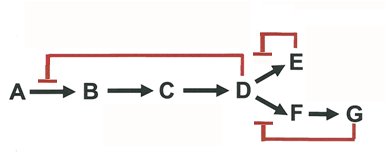
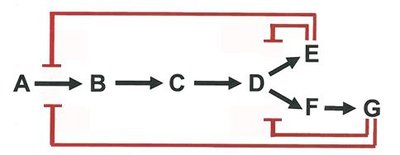
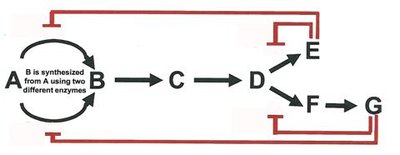
Enzyme activity is regulated through various mechanisms, including feedback inhibition, allosteric regulation, and covalent modification. Feedback inhibition is a common regulatory scheme in metabolic pathways, where the end product inhibits an early enzyme in the pathway.
Coenzymes and Essential Ions
Classification and Function
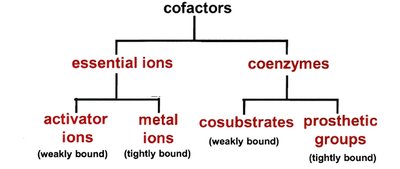
Many enzymes require non-protein cofactors for activity. These include essential ions (metal ions) and coenzymes (organic molecules). Cofactors can be tightly or loosely bound to the enzyme.
Coenzyme | Major metabolic roles | Mechanistic role |
|---|---|---|
ATP | Transfer of phosphoryl or nucleotidyl groups | Cosubstrate |
NAD+/NADP+ | Oxidation-reduction reactions | Cosubstrate |
FMN/FAD | Oxidation-reduction reactions | Prosthetic group |
Coenzyme A | Transfer of acyl groups | Cosubstrate |
Pyridoxal phosphate (PLP) | Transfer of groups to and from amino acids | Prosthetic group |
Tetrahydrofolate | Transfer of one-carbon substituents | Cosubstrate |
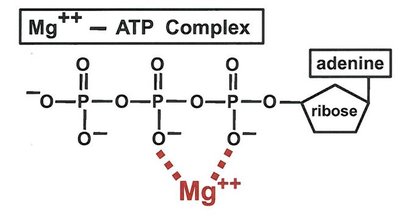
Essential Ion Cofactors
Metal ions such as Mg2+, K+, and Ca2+ are required by many enzymes for structural or catalytic roles. For example, Mg2+ is essential for ATP binding in kinases, reducing the negative charge and facilitating nucleophilic attack.
Carbohydrate Structure and Stereochemistry
Classification of Sugars
Carbohydrates are classified based on the functional group (aldose or ketose), number of carbons, and stereochemistry. Monosaccharides are the simplest carbohydrates, while oligo- and polysaccharides are formed by glycosidic linkages between monosaccharide units.
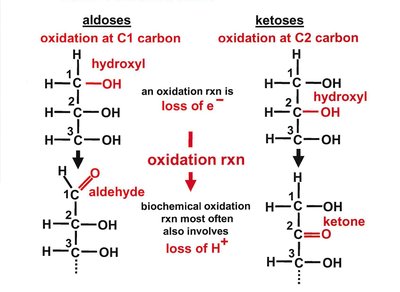
Stereochemistry of Monosaccharides
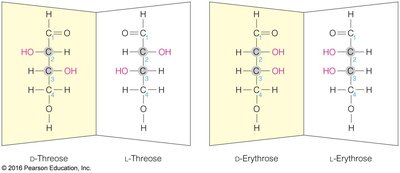
Monosaccharides exhibit chirality, with D- and L-forms based on the configuration of the chiral carbon farthest from the carbonyl group. Enantiomers are mirror images, while diastereomers differ at one or more chiral centers but are not mirror images.
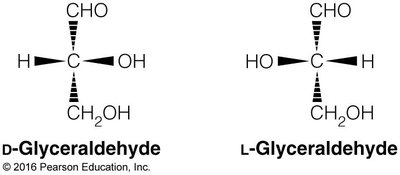
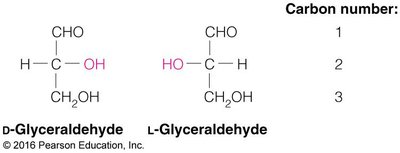
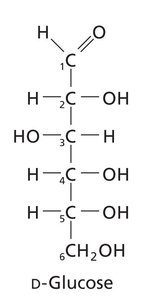
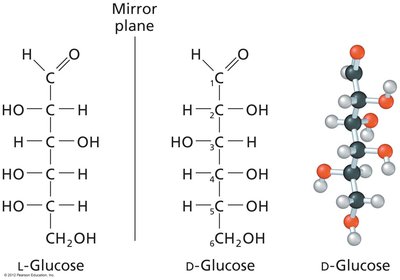
Cyclization and Anomeric Forms
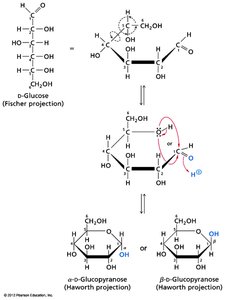
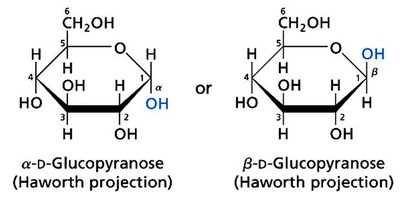
Monosaccharides can cyclize to form hemiacetals or hemiketals, generating a new chiral center at the anomeric carbon. The two possible configurations are designated as α or β anomers.
Summary Table: Polysaccharide Types and Linkages
Polysaccharide | Component | Linkages |
|---|---|---|
Amylose (starch) | Glc | α(1→4) |
Amylopectin (starch) | Glc | α(1→4), α(1→6) (branches) |
Glycogen | Glc | α(1→4), α(1→6) (branches) |
Cellulose | Glc | β(1→4) |
Chitin | GlcNAc | β(1→4) |
Hyaluronic acid | GlcUA and GlcNAc | β(1→3), β(1→4) |
Additional info: This guide covers the essential concepts of enzyme catalysis, inhibition, cofactor function, and carbohydrate structure, providing a foundation for further study in biochemistry.
