 Back
BackCell Adhesions, Cell Junctions, and Extracellular Structures: Study Guide
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
Beyond the Cell: Cell Adhesions, Cell Junctions, and Extracellular Structures
Introduction to Cell Adhesions and Extracellular Structures
Multicellular organisms rely on specialized structures to organize cells into tissues, enabling complex biological functions. These structures include cell-cell junctions and the extracellular matrix (ECM), which provide mechanical support, facilitate communication, and regulate cellular behavior.
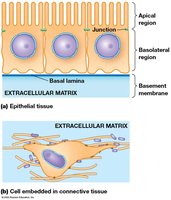
Types of Tissues in Animals
Epithelium and Connective Tissue
Animal tissues are primarily classified as epithelial or connective tissue:
Epithelium: Sheets of polarized cells with distinct apical and basal domains, forming barriers and interfaces.
Connective tissue: Loosely organized cells embedded in ECM, providing structural support and flexibility.
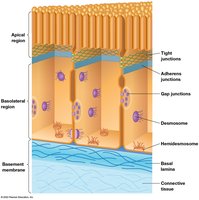
Cell-Cell Junctions
Overview of Junction Types
Cell-cell junctions are specialized structures that join cells together, forming tissues and organs. Animal cells possess three main types of junctions:
Adhesive junctions: Anchor the cytoskeleton to the cell surface.
Tight junctions: Seal cells together, preventing passage of molecules between them.
Gap junctions: Allow direct communication between cells via small channels.
Plant cells have plasmodesmata, which functionally resemble gap junctions.
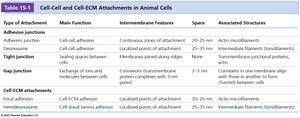
Summary Table: Cell-Cell and Cell-ECM Attachments in Animal Cells
The following table summarizes the main types of cell junctions and their features:
Type of Attachment | Main Function | Intermembrane Features | Space | Associated Structures |
|---|---|---|---|---|
Adherens junction | Cell-cell adhesion | Continuous zones of attachment | 20–25 nm | Actin microfilaments |
Desmosome | Cell-cell adhesion | Membrane paired along ridges | 25–35 nm | Intermediate filaments (keratin/desmin) |
Tight junction | Sealing spaces between cells | Membranes joined along ridges | 0 nm | Actin microfilaments |
Gap junction | Exchange of ions and molecules between cells | Connexon protein complexes with 3 nm channels | 2–3 nm | None |
Focal adhesion | Cell-ECM adhesion | Local points of attachment | 20–25 nm | Actin microfilaments |
Hemidesmosome | Cell-basal lamina adhesion | Localized points of attachment | 25–35 nm | Intermediate filaments (keratin) |
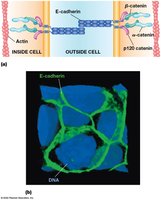
Adhesive Junctions
Adherens Junctions
Adherens junctions are cadherin-mediated structures that connect actin filaments between cells, especially in epithelial tissues. Cadherins are transmembrane proteins with extracellular repeats, a transmembrane domain, and variable cytosolic ends.
E-cadherin: The best-characterized cadherin, forms pairs via five extracellular repeats, interacting with β-catenin, α-catenin, and actin filaments.
Tissue specificity: Different cadherins are expressed in specific tissues, e.g., E-cadherin in epithelium, P-cadherin in placenta.
Desmosomes
Desmosomes are button-like points of strong adhesion, abundant in tissues subject to mechanical stress (skin, heart muscle). They use desmocollins and desmogleins (desmosomal cadherins) and linker proteins (e.g., β-catenin) to connect to intermediate filaments.
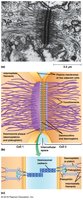
Tight Junctions
Structure and Function
Tight junctions seal epithelial cells together, forming a continuous belt around the apical ends. They contain occludin, junctional adhesion molecules (JAMs), and claudins, which interlock to form ion-selective pores for paracellular transport.
Paracellular transport: Movement of ions between cells, regulated by claudins.
Barrier function: Prevents lateral movement of membrane proteins and lipids.
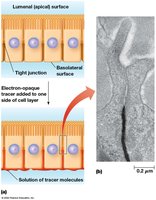
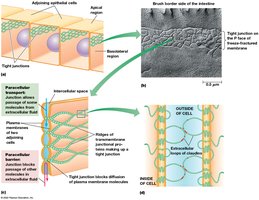
Gap Junctions
Structure and Communication
Gap junctions are regions where plasma membranes are closely aligned, joined by connexons (assemblies of six connexin proteins). They allow passage of small molecules and ions, enabling direct electrical and chemical communication between cells.
Connexon channel: ~3 nm wide, excludes proteins and nucleic acids.
Tissue specificity: Different tissues express different connexins.
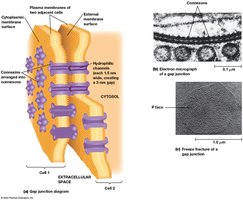
The Extracellular Matrix (ECM) of Animal Cells
ECM Composition and Function
The ECM provides structural support and regulates cell behavior. Its composition varies by tissue:
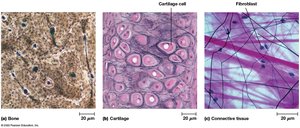
Bone: Rigid matrix with few cells.
Cartilage: Flexible matrix with abundant ECM.
Connective tissue: Gelatinous matrix with fibroblasts.
Classes of ECM Molecules
Structural proteins: Collagens and elastins (strength and flexibility).
Proteoglycans: Protein-polysaccharide complexes (hydrated matrix).
Adhesive glycoproteins: Fibronectins and laminins (cell attachment).
Collagen Structure and Assembly
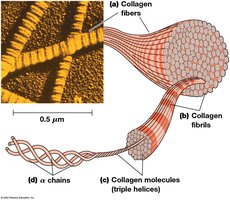
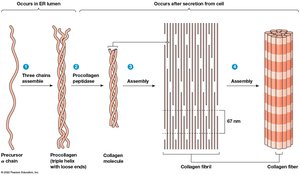
Collagen is the most abundant ECM protein, forming fibers with high tensile strength. Collagen molecules are composed of three α chains twisted into a helix, assembled into fibrils and fibers.
Elastins and ECM Elasticity
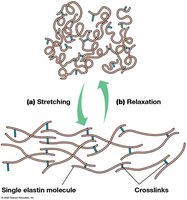
Elastins provide elasticity to the ECM, forming stretchable fibers crosslinked by lysine residues. Tension stretches the network, while relaxation returns it to its original state.
Proteoglycans and Glycosaminoglycans (GAGs)
Proteoglycans are glycoproteins with numerous GAG chains, forming a gel-like network in the ECM. Common GAGs include chondroitin sulfate, keratan sulfate, and hyaluronate.
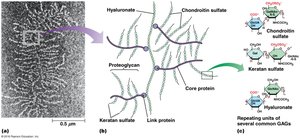
Adhesive Glycoproteins
Fibronectins
Fibronectins are glycoproteins that bind cells to the ECM and guide cellular movement. They have multiple domains for binding ECM molecules and cell surface receptors via the RGD sequence.
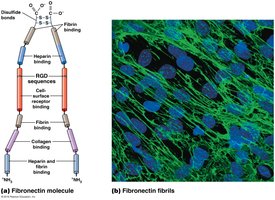
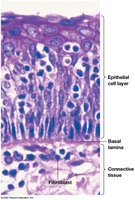
Laminins and the Basal Lamina
Laminins are found in the basal lamina, a specialized ECM layer underlying epithelial cells. The basal lamina provides structural support and acts as a permeability barrier, containing type IV collagen, proteoglycans, laminins, and entactin/nidogen.
Integrins: Cell Surface Receptors for ECM
Integrin Structure and Function
Integrins are transmembrane receptors that bind ECM constituents (fibronectins, laminins) and integrate the cytoskeleton with the ECM. Each integrin is a heterodimer of α and β subunits, recognizing RGD sequences in ECM glycoproteins.
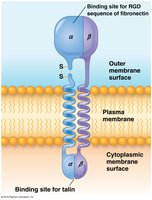
Integrins and Cytoskeletal Linkage
Integrin tails interact with cytosolic proteins (talin, vinculin, α-actinin) to link to actin filaments. Migratory and non-epithelial cells attach to ECM via focal adhesions, while epithelial cells use hemidesmosomes (α6β4 integrin linked to keratin).
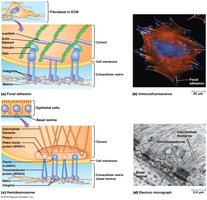
Specialized ECM Attachments
Dystrophin/Dystroglycan Complex
The dystrophin/dystroglycan complex stabilizes muscle cell attachments to the ECM, especially at costameres in striated muscle. Defects in this complex are linked to muscular dystrophy.

Plant Cell Junctions and Walls
Plasmodesmata
Plasmodesmata are cytoplasmic channels through plant cell walls, lined with plasma membrane and containing a central desmotubule. They allow direct communication between plant cells, similar to gap junctions in animals.
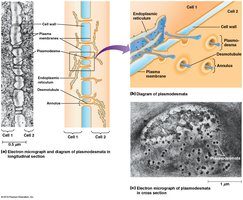
Plant Cell Wall Structure
Plant cell walls consist of cellulose microfibrils, branched polysaccharides (pectins, hemicelluloses), and glycoproteins (extensins). The wall provides structural support, withstands turgor pressure, and acts as a permeability barrier for large molecules.
Review Questions and Applications
Collagen Structure Hierarchy
From largest to smallest: collagen fiber → collagen fibril → collagen molecule → procollagen → α chain polypeptide.
Functional Analogy
Plasmodesmata in plants are most similar in function to gap junctions in animal cells, as both permit direct cell-cell communication.
Epithelial-Mesenchymal Transition (EMT)
EMT is the breakdown of epithelium into mesenchyme, accompanied by changes in cadherin expression. This process is important in cancer metastasis.
Integrins and Signaling
Integrins participate in intracellular signaling, including "inside-out" and "outside-in" pathways, affecting cell adhesion and growth.
Anchorage-Dependent Growth
Most cells require attachment to ECM for growth; integrin clustering activates intracellular pathways necessary for cell division.
Summary
Cell adhesions, junctions, and extracellular structures are fundamental to tissue organization, communication, and function in multicellular organisms. Understanding these systems is essential for cell biology, tissue engineering, and pathology.
