 Back
Back18: Protein Sorting and Intracellular Membrane Trafficking
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
Protein Sorting and Intracellular Membrane Trafficking
Introduction to Protein Sorting
Protein sorting is a fundamental process in cell biology that ensures proteins are delivered to their correct cellular destinations. This process relies on specific signal sequences within the protein structure, which direct the protein to the appropriate organelle or membrane compartment.
Signal sequences are short stretches of amino acids that act as 'addresses' for protein targeting.
Proteins without signal sequences remain in the cytosol, while those with specific signals are transported to organelles such as the endoplasmic reticulum (ER), mitochondria, chloroplasts, or peroxisomes.
Correct protein localization is essential for cellular function and viability.
Transport of Proteins Across the Endoplasmic Reticulum (ER)
Entry Points and Protein Destinations
The ER serves as the entry point for proteins destined for the secretory pathway, including the Golgi apparatus, endosomes, lysosomes, and the plasma membrane.
Water-soluble proteins are translocated across the ER membrane into the lumen, where they may be secreted or reside within organelles.
Transmembrane proteins are inserted into the ER membrane and may remain there or be transported to other membranes.
Signal Sequences: Structure and Function
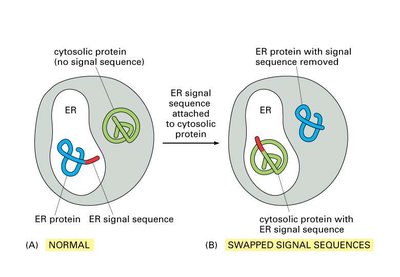
Signal sequences are both necessary and sufficient for targeting proteins to the ER. Experimental evidence demonstrates that swapping or removing these sequences alters protein localization.
Typical ER signal sequences are hydrophobic stretches near the N-terminus of the protein.
Experimental manipulation of signal sequences can redirect proteins to different cellular compartments.

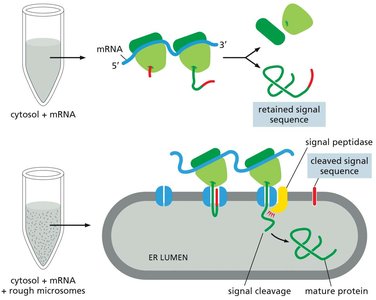
Experimental Basis for the Signal Hypothesis
The signal hypothesis was confirmed by experiments showing that proteins synthesized in the presence of ER membranes have their signal sequences cleaved, while those synthesized in their absence retain the sequence.
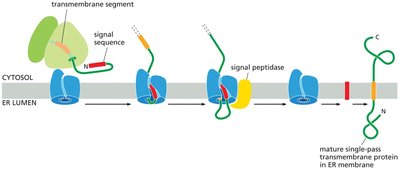
Signal peptidase in the ER membrane cleaves the signal sequence during translocation.
This process is essential for the maturation and proper localization of secretory proteins.
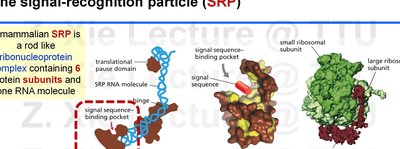
The Signal Recognition Particle (SRP) and Targeting to the ER
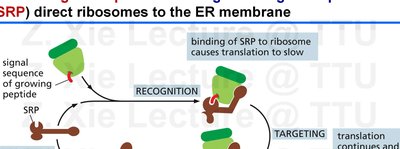
The SRP is a ribonucleoprotein complex that recognizes and binds to the ER signal sequence as it emerges from the ribosome, pausing translation and targeting the ribosome-protein complex to the ER membrane.
SRP binding causes a temporary pause in translation.
The SRP-ribosome complex is directed to the SRP receptor on the ER membrane, where translation resumes and the protein is translocated into the ER.
Free and Membrane-Bound Ribosomes
Ribosomes can be free in the cytosol or bound to the ER membrane, depending on the presence of an ER signal sequence in the nascent polypeptide.
Membrane-bound ribosomes synthesize proteins destined for the ER, Golgi, lysosomes, plasma membrane, or secretion.
Free ribosomes synthesize proteins that remain in the cytosol or are targeted to organelles such as the nucleus, mitochondria, or peroxisomes.

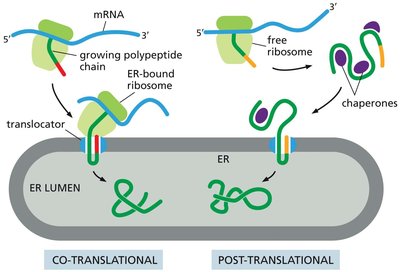
Co-Translational and Post-Translational Translocation
Proteins can be translocated into the ER either during their synthesis (co-translational) or after translation is complete (post-translational).
Co-translational translocation is the predominant mechanism in higher eukaryotes.
Post-translational translocation is more common in yeast and involves chaperone proteins to maintain the unfolded state of the protein.
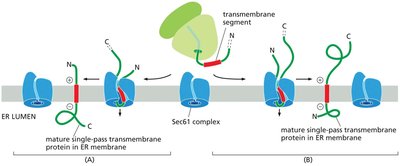
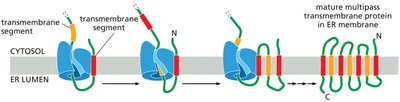
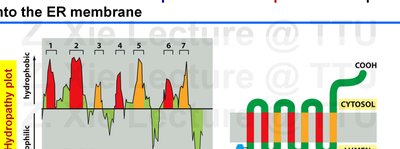
Insertion of Transmembrane Proteins into the ER Membrane
Single-Pass and Multi-Pass Transmembrane Proteins
Transmembrane proteins are integrated into the ER membrane through the action of translocon complexes, guided by hydrophobic transmembrane segments and signal sequences.
Single-pass transmembrane proteins contain one membrane-spanning segment.
Multi-pass transmembrane proteins contain multiple membrane-spanning segments, each acting as a start- or stop-transfer sequence.
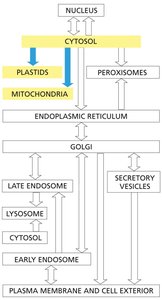
Protein Import into Mitochondria, Chloroplasts, and Peroxisomes
General Features of Organelle Targeting
Proteins destined for mitochondria, chloroplasts, and peroxisomes are synthesized in the cytosol and contain organelle-specific targeting sequences.
These proteins lack ER signal sequences and are imported post-translationally.
Targeting sequences are recognized by receptors on the organelle surface, facilitating import.
Mitochondrial Protein Import
Mitochondrial proteins contain N-terminal signal sequences that form amphipathic helices, recognized by import receptors. Translocation across mitochondrial membranes is mediated by multimeric protein complexes.
TOM (Translocator of the Outer Membrane) and TIM (Translocator of the Inner Membrane) complexes facilitate protein import.
Signal sequences are cleaved after import, and chaperones assist in protein folding.
Chloroplast Protein Import
Chloroplast precursor proteins contain an N-terminal chloroplast signal sequence, often followed by a thylakoid signal sequence. Import involves sequential translocation across the chloroplast envelope and thylakoid membrane.
Signal sequences are cleaved after import into the stroma and thylakoid.
Multiple pathways exist for translocation into the thylakoid space or membrane.
Peroxisomal Protein Import
Proteins targeted to peroxisomes contain either a C-terminal PTS1 signal or, less commonly, an N-terminal PTS2 signal. Import is mediated by cytosolic receptors and does not require protein unfolding.
Signal sequences are not removed after import.
Folded proteins can be imported into peroxisomes, distinguishing this pathway from mitochondrial and chloroplast import.
Vesicular Transport and Membrane Trafficking
Overview of Vesicular Transport
Vesicular transport is essential for the movement of proteins and lipids between membrane-bound compartments in the secretory and endocytic pathways.
Transport vesicles bud from donor compartments and fuse with target compartments, preserving membrane orientation.
Soluble components are transferred from lumen to lumen, while membrane proteins and lipids retain their orientation.
Determinants of Trafficking Specificity
Specificity in vesicular trafficking is determined by transport signals, vesicle coat proteins, and proteins involved in vesicle tethering, docking, and fusion.
Coat proteins such as clathrin, COPI, and COPII select cargo and shape vesicles.
Adaptor proteins link coat proteins to cargo receptors.
Vesicle coats are rapidly disassembled after budding to allow fusion with target membranes.
Types of Coated Vesicles
There are three main types of coated vesicles, each associated with specific trafficking steps:
Clathrin-coated vesicles: Mediate transport from the Golgi apparatus and plasma membrane.
COPI-coated vesicles: Mediate retrograde transport from the Golgi to the ER and between Golgi cisternae.
COPII-coated vesicles: Mediate anterograde transport from the ER to the Golgi apparatus.
Summary Table: Typical Signal Sequences for Protein Targeting
Destination | Signal Sequence |
|---|---|
Nucleus (import) | Pro-Pro-Lys-Lys-Lys-Arg-Lys-Val |
Nucleus (export) | Met-Glu-Glu-Leu-Ser-Gln-Ala-Leu-Ala-Ser-Ser-Phe |
Mitochondria | Met-Leu-Ser-Leu-Arg-Gln-Ser-Ile-Arg-Phe-Phe-Lys-Pro-Ala-Thr-Arg-Thr-Leu |
Plastid | Met-Val-Ala-Met-Ala-Met-Ala-Ser-Leu-Gln-Ser-Ser-Ser-Ser-Leu-Ser-Leu |
Peroxisome | Ser-Lys-Leu (C-terminal) |
ER | Met-Met-Ser-Phe-Val-Ser-Leu-Leu-Leu-Val-Gly-Ile-Leu-Phe-Trp-Ala-Thr |
Return to ER | Lys-Asp-Glu-Leu (C-terminal) |
