 Back
BackTrafficking and Sorting in the Endomembrane System: Protein Transport, Vesicle Formation, and Cellular Digestion
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
Trafficking Through the Endomembrane System
Overview of Trafficking
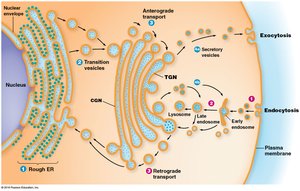
The endomembrane system is responsible for the synthesis, sorting, and transport of proteins and lipids within eukaryotic cells. Proteins released into the endoplasmic reticulum (ER) lumen are routed to the Golgi complex, secretory vesicles, or back to the ER. Sorting of proteins begins in the ER and early Golgi compartments, with mechanisms to retrieve or retain compartment-specific proteins. The final sorting of material that will leave the Golgi complex occurs in the trans-Golgi network (TGN).
Key Point 1: Proteins are sorted and trafficked from the ER to the Golgi, secretory vesicles, or back to the ER.
Key Point 2: Sorting mechanisms include retention and retrieval tags, and final sorting occurs in the TGN.
Example: Secretory proteins are routed through the Golgi to vesicles for exocytosis.
Protein Tags: Retention and Retrieval
Protein tags are molecular features that direct proteins to specific compartments. Tags may be amino acid sequences, hydrophobic domains, or oligosaccharide side chains. ER-specific proteins contain retention tags (e.g., RXR) and retrieval tags (e.g., KDEL, KKXX in mammals; HDEL in yeast) to ensure proper localization.
Key Point 1: Retention tags prevent proteins from escaping the ER; retrieval tags return proteins from the Golgi to the ER.
Key Point 2: Tags bind to specific receptors, which package the protein into transport vesicles for return.
Example: KDEL-tagged proteins are retrieved from the Golgi to the ER.
Sorting of Golgi Complex Proteins
Golgi-specific proteins are integral membrane proteins, and their sorting may depend on the length of their membrane-spanning domains. The thickness of cellular membranes increases from the ER to the plasma membrane, and proteins move until the membrane thickness exceeds their transmembrane domain length.
Key Point 1: Membrane thickness increases from ER (5 nm) to plasma membrane (8 nm).
Key Point 2: Proteins are sorted based on the compatibility of their transmembrane domain length with membrane thickness.
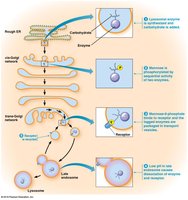
Targeting of Soluble Lysosomal Proteins
Soluble lysosomal enzymes are synthesized in the ER and undergo N-glycosylation, followed by phosphorylation of mannose residues in the Golgi, forming mannose-6-phosphate tags. These tags ensure delivery of lysosomal proteins to lysosomes via receptor-mediated sorting in the TGN.
Key Point 1: Mannose-6-phosphate tags direct lysosomal enzymes to lysosomes.
Key Point 2: Sorting occurs in the TGN, with receptor-mediated transport to endosomes and lysosomes.
Example: Lysosomal hydrolases are tagged and delivered to lysosomes for cellular digestion.
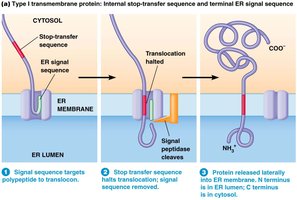
Insertion of Integral Membrane Proteins into the ER
Stop-Transfer Sequences
Integral membrane proteins are synthesized with ER signal sequences. The stop-transfer sequence, a hydrophobic domain, halts translocation through the ER membrane, resulting in a permanent transmembrane segment. The signal sequence is cleaved, and the protein is released laterally into the membrane.
Key Point 1: Stop-transfer sequences halt translocation and anchor proteins in the ER membrane.
Key Point 2: The N-terminus is in the ER lumen, and the C-terminus is in the cytosol.
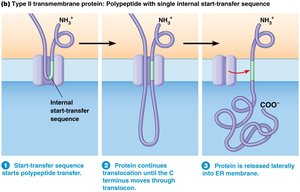
Internal Start-Transfer Sequences
Some membrane proteins lack a typical N-terminal signal sequence and instead have internal start-transfer sequences. These sequences are recognized by SRP and targeted to the ER membrane, where the hydrophobic region functions as a membrane anchor.
Key Point 1: Internal start-transfer sequences initiate translocation and anchor proteins in the membrane.
Key Point 2: The C-terminus is released into the ER membrane.
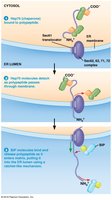
Posttranslational Import into the ER Lumen
Some proteins are synthesized in the cytosol and imported into the ER lumen posttranslationally. Chaperones (Hsp70 family) keep the protein unfolded, and the Sec61 pore complex targets the protein for translocation. BiP couples ATP hydrolysis to pull the polypeptide into the ER lumen.
Key Point 1: Posttranslational import uses chaperones and the Sec61 pore for translocation.
Key Point 2: BiP and ATP hydrolysis drive the import process.
Transport Across the Plasma Membrane: Exocytosis and Endocytosis
Exocytosis
Exocytosis is the process by which secretory vesicles release their contents outside the cell. Secretory pathways move proteins from the ER through the Golgi to secretory vesicles, which discharge their contents to the exterior. Constitutive secretion is continuous and unregulated, while regulated secretion occurs in response to specific signals.
Key Point 1: Exocytosis involves vesicle fusion with the plasma membrane and release of contents.
Key Point 2: Constitutive secretion is continuous; regulated secretion is triggered by signals (e.g., neurotransmitter release).
Example: Mucus secretion by intestinal lining (constitutive); neurotransmitter release (regulated).
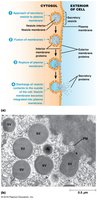
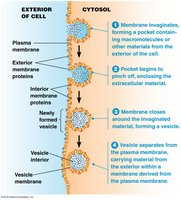
Endocytosis
Endocytosis is the process by which cells internalize external materials by forming vesicles from the plasma membrane. Endocytic vesicles develop into early endosomes, which fuse with vesicles from the TGN and acquire digestive enzymes to form new lysosomes. Phagocytosis ingests solid particles, while pinocytosis takes up liquids.
Key Point 1: Endocytosis forms vesicles from the plasma membrane to internalize materials.
Key Point 2: Endocytic vesicles mature into endosomes and lysosomes.
Example: Phagocytosis of bacteria; pinocytosis of extracellular fluid.
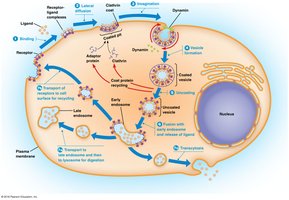
Receptor-Mediated Endocytosis
Receptor-mediated endocytosis (clathrin-dependent) allows cells to internalize specific macromolecules using cell surface receptors. Ligands bind to receptors, which accumulate in coated pits. Adaptor proteins, clathrin, and dynamin induce curvature and invagination, forming coated vesicles that fuse with early endosomes.
Key Point 1: Receptor-ligand complexes are internalized via coated pits and vesicles.
Key Point 2: Clathrin, adaptor proteins, and dynamin drive vesicle formation and separation.
Example: Uptake of cholesterol, hormones, and growth factors.
Coated Vesicles and Vesicular Transport
Clathrin-Coated Vesicles
Clathrin-coated vesicles are surrounded by lattices composed of clathrin and adaptor proteins. The basic unit of clathrin lattices is a triskelion, which assembles into hexagons and pentagons to close the coat and curve around the budding vesicle. The assembly of the clathrin coat provides the driving force for vesicle formation.
Key Point 1: Clathrin triskelions assemble into lattices for vesicle formation.
Key Point 2: Adaptor proteins and dynamin are required for vesicle constriction and separation.
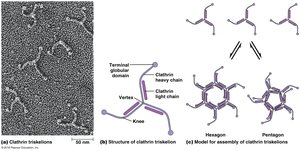

COPI- and COPII-Coated Vesicles
COPI-coated vesicles mediate retrograde transport from the Golgi back to the ER, while COPII-coated vesicles mediate anterograde transport from the ER to the Golgi. Coat assembly is mediated by small GTP-binding proteins (ARF for COPI, SarI for COPII), and requires energy from GTP hydrolysis.
Key Point 1: COPI vesicles transport proteins from Golgi to ER; COPII vesicles transport from ER to Golgi.
Key Point 2: Coat assembly is regulated by GTP-binding proteins.
Coated Vesicle | Coat Proteins | Origin | Destination |
|---|---|---|---|
Clathrin | Clathrin, AP1, ARF | TGN | Endosomes |
Clathrin | Clathrin, AP2 | Plasma membrane | Endosomes |
COPI | COPI, ARF | Golgi apparatus | ER or Golgi apparatus |
COPII | COPII (Sec13/31 and Sec23/24), Sar1 | ER | Golgi apparatus |
Caveolin | Caveolin | Plasma membrane | ER? |
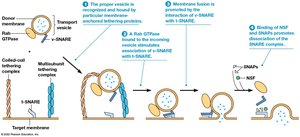
SNARE Proteins and Vesicle Fusion
SNARE-Mediated Fusion
SNARE proteins mediate fusion between vesicles and target membranes. v-SNAREs are found on vesicles, and t-SNAREs are found on target membranes. Rab GTPases promote binding, and NSF/SNAPs dissolve the SNARE complex after fusion. Tethering proteins interact with incoming vesicles over longer distances.
Key Point 1: v-SNAREs and t-SNAREs allow recognition and fusion of vesicles with target membranes.
Key Point 2: Rab GTPases, NSF, SNAPs, and tethering proteins regulate the fusion process.
Lysosomes and Cellular Digestion
Lysosome Structure and Function
Lysosomes are organelles containing digestive enzymes capable of degrading all major classes of biological macromolecules. Lysosomes maintain an acidic environment (pH 4.0–5.0) via ATP-dependent proton pumps (V-ATPases). Lysosomal enzymes are acid hydrolases, and lysosomes develop from endosomes as they acquire these enzymes.
Key Point 1: Lysosomes are highly acidic and contain acid hydrolases for digestion.
Key Point 2: Lysosomal enzymes are delivered from the TGN to endosomes, which mature into lysosomes.
Lysosomal Storage Diseases
Lysosomal storage diseases are characterized by the accumulation of indigestible material due to absent lysosomal proteins. Examples include Type II glycogenosis (excessive glycogen), Hurler and Hunter syndromes (glycosaminoglycans), and Tay-Sachs disease (ganglioside accumulation).
Key Point 1: Over 40 lysosomal storage diseases are known, most are not treatable.
Key Point 2: Accumulation of undigested substances leads to cellular dysfunction.
Example: Tay-Sachs disease affects the nervous system due to ganglioside accumulation.
Peroxisomes
Structure and Function
Peroxisomes are single-membrane-bound organelles characterized by the presence of catalase for degrading hydrogen peroxide (H2O2). They are involved in hydrogen peroxide metabolism, detoxification, oxidation of fatty acids (β-oxidation), metabolism of nitrogen-containing compounds, and catabolism of unusual substances.
Key Point 1: Catalase degrades H2O2 to protect cells from oxidative damage.
Key Point 2: Peroxisomes play roles in detoxification and fatty acid metabolism.
Autophagy and Cellular Recycling
Autophagy and Cancer
Autophagy is the process by which cells degrade damaged or unneeded cellular structures. Recent research suggests a direct link between autophagy and cancer, as defective autophagy can contribute to tumorigenesis.
Key Point 1: Autophagy is essential for cellular homeostasis and recycling.
Key Point 2: Impaired autophagy is associated with cancer development.
Summary Table: Key Sorting Tags and Mechanisms
Tag/Mechanism | Function | Example |
|---|---|---|
RXR | ER retention | NMDA receptor subunits |
KDEL/KKXX | ER retrieval | Soluble ER proteins |
Mannose-6-phosphate | Lysosomal targeting | Lysosomal hydrolases |
Additional info: These notes expand on the original content with academic context, definitions, and examples to ensure completeness and clarity for cell biology students.
