 Back
BackTransport of Water, Minerals, and Sugars in Plants
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
Absorption and Transport of Water and Minerals in Plants
Absorption by Root Cells
Water and mineral absorption in plants primarily occurs through root hairs located near the root tips. The extensive surface area of cortical cell membranes enhances the uptake of water and selected minerals from the soil. The concentration of essential minerals is typically higher in the roots than in the surrounding soil due to active transport mechanisms.
Root hairs increase the surface area for absorption.
Active transport allows roots to accumulate minerals against their concentration gradient.
Soil solution enters the root and moves toward the vascular tissue.
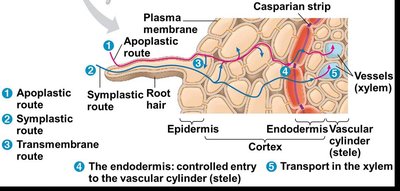
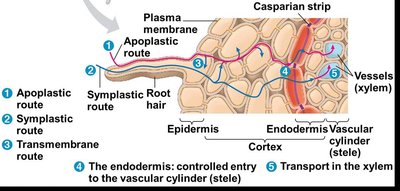
Pathways for Water and Mineral Movement
Once inside the root, water and minerals can move toward the vascular cylinder (stele) via three main pathways:
Apoplastic route: Movement along cell walls and extracellular spaces without crossing cell membranes.
Symplastic route: Movement through the cytoplasm of cells interconnected by plasmodesmata.
Transmembrane route: Movement from cell to cell by repeatedly crossing plasma membranes and cell walls.
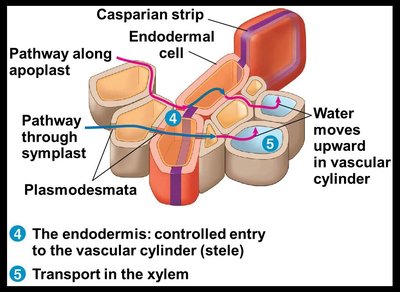
Role of the Endodermis and Casparian Strip
The endodermis is the innermost layer of the root cortex and acts as a selective barrier. The Casparian strip, a band of suberin in the endodermal cell walls, blocks the apoplastic route, forcing water and minerals to cross a selectively permeable plasma membrane before entering the vascular cylinder (xylem).
Ensures selective uptake of minerals.
Prevents backflow of solutes from the stele to the cortex.
Bulk Flow Transport via the Xylem
Mechanism of Bulk Flow
Xylem sap, consisting of water and dissolved minerals, is transported from roots to leaves by bulk flow. This process is driven by transpiration—the loss of water vapor from the plant's surface, primarily through stomata. As water evaporates from the leaves, it creates a negative pressure that pulls water upward from the roots through the xylem.
Bulk flow is much faster than diffusion and moves the entire solution.
Driven by differences in pressure potential, not solute potential.
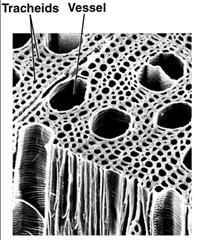
Occurs in hollow, dead xylem cells (tracheids and vessel elements).

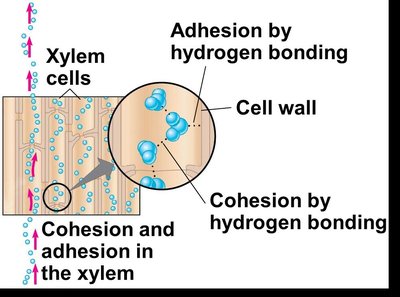
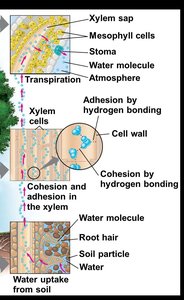
The Cohesion-Tension Hypothesis
The cohesion-tension hypothesis explains how water is pulled upward through the plant. Transpiration at the leaf surface creates a negative pressure (tension) in the xylem, which is transmitted down to the roots. Water molecules stick together (cohesion) and to the walls of xylem cells (adhesion), forming a continuous water column.
Cohesion: Hydrogen bonding between water molecules.
Adhesion: Hydrogen bonding between water molecules and cell walls.
Transpirational pull: Generated as water vapor exits the leaf, pulling the water column upward.
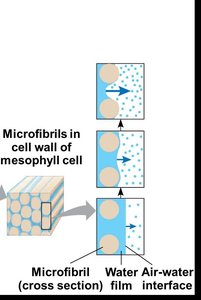
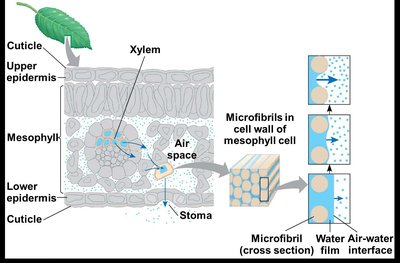
Water Potential Gradient
Water moves from regions of higher water potential (soil) to lower water potential (atmosphere). The water potential gradient drives the upward movement of water through the plant.
Water potential () is measured in megapascals (MPa).
Typical values: Soil MPa, Leaf air spaces MPa, Outside air MPa.
Adaptations to Prevent Xylem Collapse
Thick secondary walls in vessel elements and tracheids prevent collapse under tension. Drought or freezing can cause cavitation (blockage of water channels), but plants can detour water through pits or use new xylem layers.
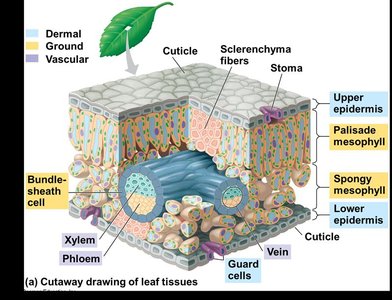
Regulation of Transpiration by Stomata
Structure and Function of Stomata
Stomata are pores on the leaf surface that regulate gas exchange and water loss. Each stoma is flanked by two guard cells that control its opening and closing by changing shape in response to turgor pressure.
Stomatal density is influenced by genetic and environmental factors.
About 95% of water loss in plants occurs through stomata.
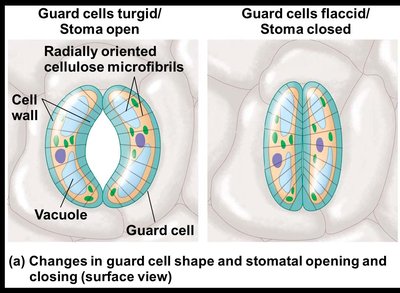
Mechanisms of Stomatal Opening and Closing
Changes in turgor pressure, primarily due to the reversible uptake and loss of potassium ions (K+), cause guard cells to bow outward (opening the stoma) or become less bowed (closing the stoma).
When guard cells are turgid, the stoma opens.
When guard cells are flaccid, the stoma closes.
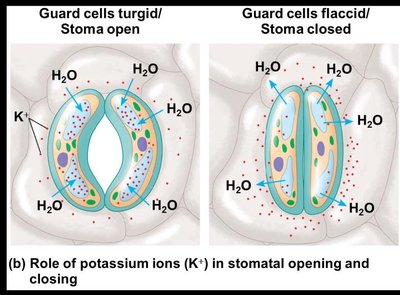
Stimuli for Stomatal Movement
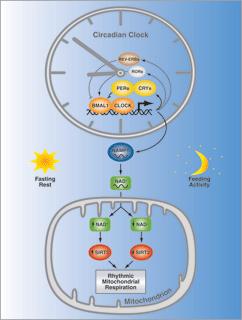
Stomata generally open during the day and close at night to minimize water loss. Opening is triggered by light, CO2 depletion, and an internal circadian clock in guard cells.
All eukaryotic organisms have internal clocks (circadian rhythms).
Adaptations to Reduce Water Loss
Xerophytes are plants adapted to arid climates. Adaptations include thick cuticles, sunken stomata, reduced leaves, and short life cycles during rainy seasons. Some use crassulacean acid metabolism (CAM) photosynthesis, opening stomata at night to reduce water loss.
Stomata recessed in crypts lined with hairs.
Leaves reduced to spines or dropped during dry periods.
CAM plants exchange gases at night.




Transport of Sugars in Plants: Phloem Translocation
Phloem Structure and Function
The products of photosynthesis (mainly sucrose) are transported through the phloem by translocation. In angiosperms, sieve-tube elements are the main conduits for this process. Phloem sap moves from sources (net producers, e.g., mature leaves) to sinks (net consumers or storers, e.g., roots, tubers).
Phloem sap is an aqueous solution high in sucrose.
Direction of flow depends on source-sink relationships, which can change seasonally.
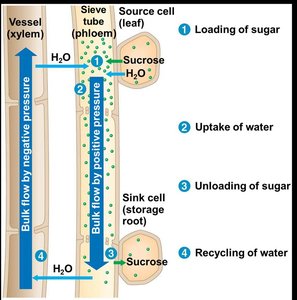
Mechanism of Phloem Loading and Unloading
Sugar is loaded into sieve-tube elements by active transport, often involving proton pumps and cotransport of sucrose and H+. At the sink, sugar diffuses into sink tissues and is followed by water, maintaining the flow.
Companion cells assist in solute movement between apoplast and symplast.
Phloem loading may be symplastic or involve both symplastic and apoplastic pathways.
Bulk Flow by Positive Pressure (Pressure Flow Hypothesis)
Phloem sap moves through sieve tubes by bulk flow driven by positive pressure, known as the pressure flow mechanism. Water uptake at the source increases pressure, driving sap toward the sink, where unloading and water loss reduce pressure.
Bulk flow in phloem is much faster than diffusion.
Direction of flow is always from source to sink.
Summary Table: Comparison of Xylem and Phloem Transport
Feature | Xylem | Phloem |
|---|---|---|
Direction of Flow | Unidirectional (roots to leaves) | Bidirectional (source to sink) |
Main Transported Substance | Water, minerals | Sugars (sucrose), amino acids |
Driving Force | Negative pressure (tension) | Positive pressure (pressure flow) |
Cell Type | Dead (tracheids, vessels) | Living (sieve-tube elements) |
