 Back
BackEnzyme Catalysis and Kinetics: Chymotrypsin as a Model
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
Enzyme Catalysis and Reaction Mechanisms
General Principles of Enzyme Catalysis
Enzymes are biological catalysts that accelerate chemical reactions by lowering the activation energy required for the reaction to proceed. They achieve this by binding reactants (substrates) in the correct orientation and stabilizing the transition state.
Activation Energy (Ea): The minimum energy required for a reaction to occur.
Transition State: A high-energy, unstable state during the reaction where old bonds are breaking and new bonds are forming.
Enzyme Mechanisms: Include nucleophilic, electrophilic, acid, and base catalysis.
Example: The rate of a reaction can be described by the Arrhenius equation:

Chymotrypsin: A Model Enzyme
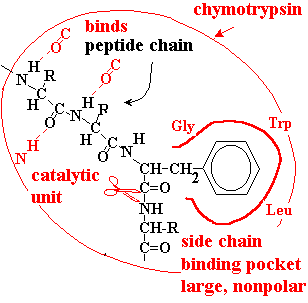
Substrate Binding and Specificity
Chymotrypsin is a protease that catalyzes the hydrolysis of peptide bonds, specifically those adjacent to aromatic amino acids (Phe, Tyr, Trp). The enzyme binds the peptide chain and positions the target bond in its catalytic site for efficient cleavage.
Binding Pocket: Large, nonpolar pocket accommodates aromatic side chains.
Specificity: Substrate fits tightly if the target amino acid matches the pocket.
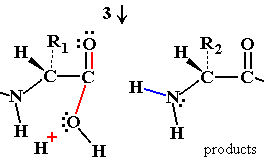
Peptide Hydrolysis Mechanism (Uncatalyzed)
Without a catalyst, water acts as a nucleophile to attack the peptide bond, but the reaction is slow due to poor nucleophilicity and an unstable transition state.
Nucleophilic Attack: Water's lone pair attacks the electron-deficient carbonyl carbon.
Transition State: Formation of an oxyanion intermediate, which is energetically unfavorable.
Bond Breakdown: The peptide bond breaks if nitrogen reclaims the excess electrons.
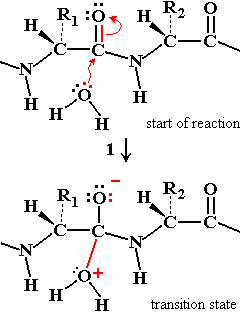
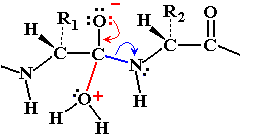
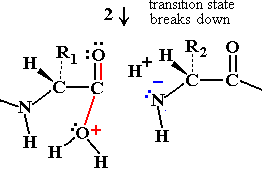
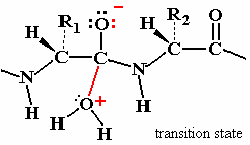
Chymotrypsin Catalytic Mechanism
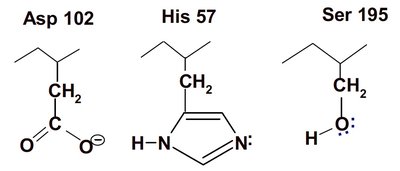
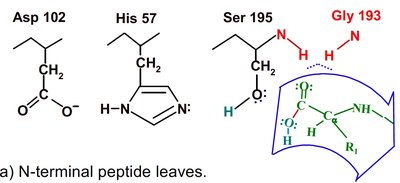
Chymotrypsin uses a two-step mechanism involving a catalytic triad (Asp 102, His 57, Ser 195) to facilitate peptide bond hydrolysis. The enzyme forms a covalent intermediate and stabilizes the transition state, greatly increasing reaction rate.
Catalytic Triad: Asp 102 (negative), His 57 (can accept H+), Ser 195 (can donate H+).
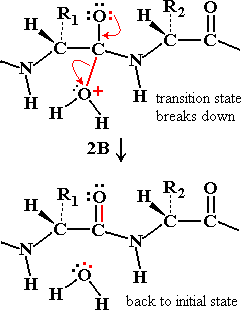
Step 1: Ser 195 acts as a nucleophile, attacking the peptide C=O, forming an acyl-enzyme intermediate.
Step 2: Water enters, His 57 activates it, and the acyl-enzyme intermediate is hydrolyzed, releasing the N-terminal peptide and regenerating the enzyme.
Transition State Stabilization
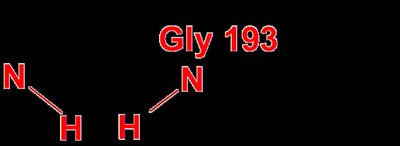
The transition state is stabilized by the oxyanion hole, which is formed by backbone N-H groups (Gly 193 and Ser 195) that provide hydrogen bonds to the negatively charged oxygen in the transition state.
Oxyanion Hole: Stabilizes the negative charge developed during the transition state.
Effect: Lowers activation energy, increases reaction rate.

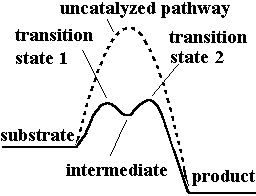
Reaction Energy Profile
Enzyme-catalyzed reactions have lower activation energy and often proceed via intermediates, as shown in the energy profile diagram.
Enzyme Assay and Detection
Measuring Enzyme Activity
Enzyme assays measure the rate of substrate disappearance or product formation. The rate is proportional to enzyme concentration and can be analyzed mathematically to determine kinetic parameters.
Enzyme Activity: Moles of substrate converted per unit time (µmol min-1).
Specific Activity: Enzyme units per milligram of total protein (µmol min-1 mg-1).
Molar Activity: Activity per mole of enzyme (min-1).
Turnover Number: Number of substrate molecules converted per enzyme molecule per second.
Spectrophotometric Assays
Many enzyme reactions are monitored by changes in absorbance using a spectrophotometer. The Beer-Lambert Law relates absorbance to concentration:
A: Absorbance
ε: Extinction coefficient (L mol-1 cm-1)
c: Concentration (mol L-1)
l: Path length (cm)
Enzyme Kinetics: Michaelis-Menten Model
Basic Kinetic Model
The Michaelis-Menten model describes how reaction rate varies with substrate concentration. The enzyme binds substrate to form an ES complex, which then produces product and releases the enzyme.
Initial Rate (v0): Measured at time zero, before product accumulates.
Vmax: Maximum rate when all enzyme is saturated with substrate.
KM: Substrate concentration at which v0 = 0.5 Vmax.

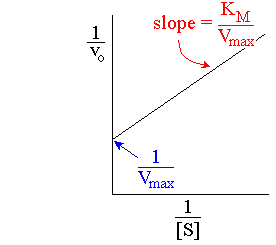
Lineweaver-Burk Plot
The Lineweaver-Burk plot linearizes the Michaelis-Menten equation by plotting 1/v0 versus 1/[S]. This allows easier determination of Vmax and KM from experimental data.
Slope: KM/Vmax
Y-intercept: 1/Vmax
X-intercept: -1/KM
Enzyme Inhibition
Types of Inhibition
Enzyme inhibitors decrease catalytic activity and are classified as competitive or non-competitive (including mixed inhibition).
Competitive Inhibition: Inhibitor binds to the active site, preventing substrate binding. Increases apparent KM, no effect on Vmax.
Non-competitive Inhibition: Inhibitor binds elsewhere, affecting catalytic activity. Decreases Vmax, no effect on KM.
Mixed Inhibition: Both Vmax and KM change.
Type | Inhibitor Binds | Effect on KM | Effect on Vmax |
|---|---|---|---|
Competitive | Active site | Increased | Unchanged |
Non-competitive | Elsewhere | Unchanged | Decreased |
Mixed | Both | Changed | Changed |
Summary
Enzymes are essential biological catalysts that increase reaction rates by lowering activation energy and stabilizing transition states. Chymotrypsin exemplifies these principles through its specific substrate binding, catalytic triad, and transition state stabilization. Enzyme kinetics, described by the Michaelis-Menten model and analyzed with Lineweaver-Burk plots, provide quantitative measures of enzyme efficiency and inhibition.
