 Back
BackMicrobial Metabolic Diversity: Catabolism, Respiration, and Energy Conservation
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
Microbial Metabolic Diversity
Overview of Microbial Catabolism
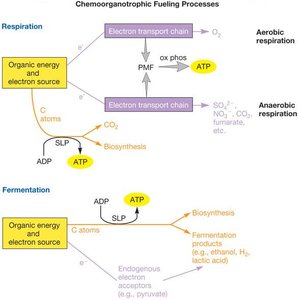
Microorganisms display remarkable metabolic diversity, allowing them to thrive in a wide range of environments. Catabolism refers to the breakdown of organic or inorganic molecules to release energy, which is then conserved as ATP. The fate of pyruvate after glycolysis depends on the availability of terminal electron acceptors and the organism's ability to detoxify oxygen radicals.
Aerobic Respiration: Oxygen is present and acts as the terminal electron acceptor.
Anaerobic Respiration: Oxygen is absent, but other electron acceptors (e.g., nitrate, sulfate) are available.
Fermentation: No external electron acceptor is present, or the organism is sensitive to oxygen.
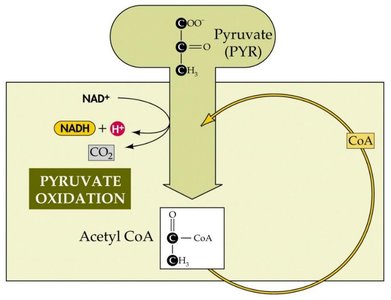
Pyruvate Oxidation and the Citric Acid Cycle (TCA/Krebs Cycle)
Pyruvate Dehydrogenase Complex
Under aerobic and anaerobic conditions, pyruvate is oxidized to acetyl-CoA by the pyruvate dehydrogenase complex. This process releases CO2 and reduces NAD+ to NADH, capturing energy for the cell.
Acetyl-CoA: Central intermediate that enters the citric acid cycle.
Energy Conservation: Some energy is conserved as NADH and in the thioester bond of acetyl-CoA.
The Citric Acid Cycle (TCA/Krebs Cycle)
The citric acid cycle is a series of enzyme-catalyzed reactions that oxidize acetyl-CoA to CO2, generating NADH, FADH2, and ATP (or GTP). It is a central metabolic hub, providing precursors for biosynthetic pathways.
Key Products per Turn: 3 NADH, 1 FADH2, 1 ATP (or GTP), 2 CO2
Precursor Metabolites: Intermediates are used for synthesis of amino acids, nucleotides, fatty acids, and porphyrins.
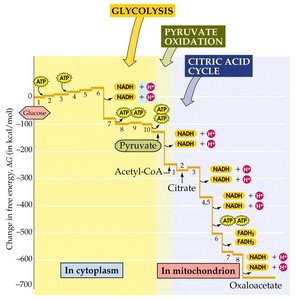
Metabolic Integration and Biosynthetic Precursors
The TCA cycle and glycolysis generate intermediates that serve as precursors for anabolic (biosynthetic) pathways, supporting cell growth and maintenance.
Fatty acids and sterols: Derived from acetyl-CoA.
Amino acids: Synthesized from TCA intermediates such as α-ketoglutarate and oxaloacetate.
Nucleic acids: Precursors from glycolysis and TCA cycle.
Porphyrin rings: Heme and chlorophyll biosynthesis.
Electron Transport Chain (ETC) and Energy Conservation
Structure and Function of the ETC
The electron transport chain is a series of protein complexes and mobile carriers that transfer electrons from donors (e.g., NADH, FADH2) to terminal electron acceptors (e.g., O2, NO3-, SO42-). This process generates a proton motive force (PMF) across the membrane, which is used to synthesize ATP.
Initial Oxidoreductase: Receives electrons from NADH or FADH2.
Mobile Electron Carriers: Quinones (e.g., ubiquinone) shuttle electrons between complexes.
Terminal Oxidoreductase: Transfers electrons to the final electron acceptor.
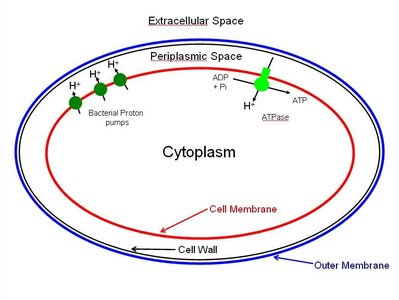
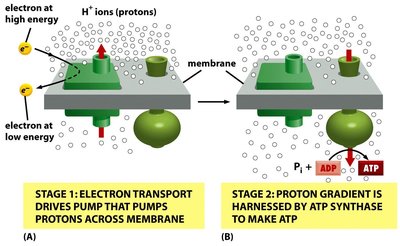
Proton Motive Force and ATP Synthesis
As electrons move through the ETC, protons are pumped across the membrane, creating an electrochemical gradient (PMF). ATP synthase uses this gradient to convert ADP and inorganic phosphate (Pi) into ATP—a process known as oxidative phosphorylation.
ATP Synthase: Enzyme complex that synthesizes ATP using the energy stored in the PMF.
Other Uses of PMF: Drives transport systems and powers flagellar rotation.
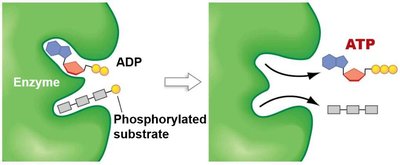
Substrate-Level vs. Oxidative Phosphorylation
Cells generate ATP by two main mechanisms:
Substrate-Level Phosphorylation (SLP): Direct transfer of a phosphate group to ADP from a phosphorylated intermediate (e.g., glycolysis, TCA cycle).
Oxidative Phosphorylation: ATP synthesis driven by the PMF generated by the ETC.

Respiratory Diversity in Prokaryotes
Aerobic vs. Anaerobic Respiration
Prokaryotes can use a variety of electron donors and acceptors, allowing them to respire in both oxic and anoxic environments. Anaerobic respiration is less energy-efficient than aerobic respiration but enables survival in the absence of oxygen.
Aerobic Respiration: O2 is the terminal electron acceptor.
Anaerobic Respiration: Alternative acceptors include NO3-, SO42-, Fe3+, etc.
Example: Escherichia coli can switch between aerobic and anaerobic respiration depending on environmental conditions.
Reversing the TCA Cycle (rTCA)
Some prokaryotes can run the TCA cycle in reverse (reductive TCA cycle) to fix CO2 and generate biosynthetic precursors, a process important for autotrophic growth.
Purpose: Removes excess NADH/FADH2, generates anabolic precursors, and fixes CO2.
Occurrence: Many bacteria and some archaea, but not in eukaryotes.
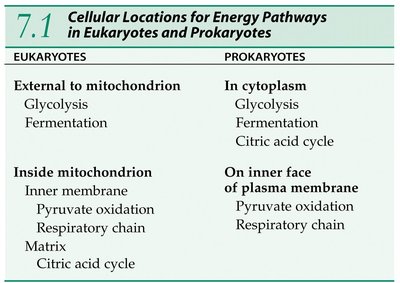
Cellular Locations of Energy Pathways
Compartmentalization in Eukaryotes and Prokaryotes
The location of metabolic pathways differs between eukaryotic and prokaryotic cells due to the presence or absence of organelles such as mitochondria.
Eukaryotes | Prokaryotes | |
|---|---|---|
Glycolysis | External to mitochondrion (cytoplasm) | In cytoplasm |
Fermentation | External to mitochondrion (cytoplasm) | In cytoplasm |
Pyruvate Oxidation | Inside mitochondrion (inner membrane) | On inner face of plasma membrane |
Respiratory Chain | Inside mitochondrion (inner membrane) | On inner face of plasma membrane |
Citric Acid Cycle | Inside mitochondrion (matrix) | In cytoplasm |
Summary Table: Electron Donors and Acceptors in Microbial Metabolism
Metabolism Type | Electron Donor | Electron Acceptor |
|---|---|---|
Chemoorganotrophy | NADH, FADH2 | O2 (aerobic), NO3-, SO42- (anaerobic) |
Chemolithotrophy | H2S, Fe2+, NH4+ | O2, NO3-, Fe3+ |
Phototrophy | H2O, H2S (excited by light) | CO2, O2 |
Key Equations
ATP Synthesis (Oxidative Phosphorylation):
General Redox Reaction:
Conclusion
Microbial metabolic diversity is rooted in the ability to utilize various catabolic pathways, electron donors, and acceptors. The integration of glycolysis, the TCA cycle, and the electron transport chain enables efficient energy conservation and biosynthesis, supporting microbial life in diverse environments.
