 Back
BackChromosome Structure, Number, and Variation in Eukaryotes
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
Chromosome Number and Structure
Species-Specific Chromosome Number and Shape
Chromosome number, size, and shape are unique to each species and are critical for understanding heredity and genetic variation. Closely related species tend to have similar chromosome numbers, but the total number can vary widely across the animal kingdom. Each chromosome pair in a diploid genome has a distinct size, shape, and genetic content, which can be visualized using cytogenetic techniques.
Genome content and chromosome number are species-specific.
Chromosome number is often conserved among closely related species.
Chromosomes can be visualized and analyzed for their size, shape, and genetic content.
Species | Diploid Chromosome Number (2n) |
|---|---|
Carp (Cyprinus carpio) | 104 |
Cat (Felis catus) | 38 |
Chicken (Gallus gallus domesticus) | 78 |
Chimpanzee (Pan troglodytes) | 48 |
Cow (Bos taurus) | 60 |
Dog (Canis familiaris) | 78 |
Frog (Rana pipiens) | 26 |
Fruit fly (Drosophila melanogaster) | 8 |
Horse (Equus caballus) | 64 |
Human (Homo sapiens) | 46 |
Mouse (Mus musculus) | 40 |
Rat (Rattus norvegicus) | 42 |
Rhesus monkey (Macaca mulatta) | 42 |
Chromosomes in the Nucleus and Chromosome Territories
Chromosomes are not randomly distributed within the nucleus. Instead, they occupy specific regions known as chromosome territories during interphase. This spatial organization is important for gene regulation and nuclear architecture.
Chromosome movement during meiosis mirrors gene transmission.
Chromosome territories are stable during interphase but dynamic during transcription and replication.
Centromeres anchor chromosomes within their territories.
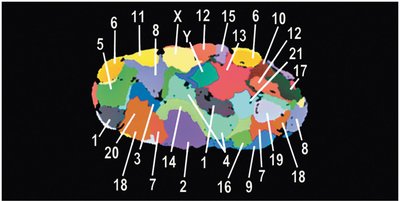
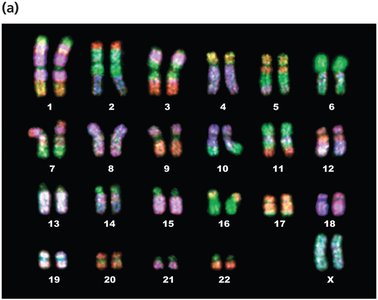
Karyotypes and Chromosome Visualization
A karyotype is an organized visual display of chromosomes, typically arranged by size and centromere position. Karyotyping is used to detect chromosomal abnormalities and to study chromosome structure.
Autosomal chromosomes are numbered 1–22; sex chromosomes are labeled X and Y.
Chromosomes can be stained to reveal unique banding patterns for identification.
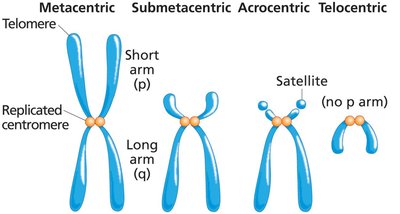
Chromosome Shape and Centromere Position
Chromosomes are classified by the position of their centromeres, which divides them into arms of unequal length. The short arm is called the p arm, and the long arm is the q arm.
Metacentric: Centromere near the middle.
Submetacentric: Centromere between the center and the tip.
Acrocentric: Centromere close to one end.
Telocentric: Centromere at the tip; no p arm.
Chromosome Banding and Molecular Cytogenetics
Chromosome Banding Techniques
Chromosome banding allows for the identification of individual chromosomes based on their unique patterns. The most common method is Giemsa (G) banding, which produces reproducible light and dark bands.
Banding patterns are used to detect structural abnormalities.
Each band can contain multiple genes.

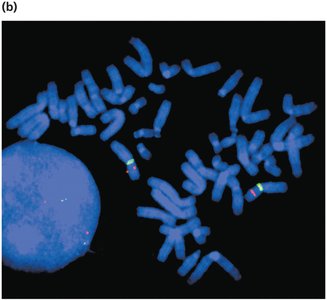
Fluorescent In Situ Hybridization (FISH)
FISH uses fluorescently labeled DNA probes to detect specific DNA sequences on chromosomes. This technique is valuable for identifying gene locations, chromosomal rearrangements, and microdeletions or duplications.
Multiple probes with different fluorescent labels can be used simultaneously.
Chromosome Number Variation: Aneuploidy and Polyploidy
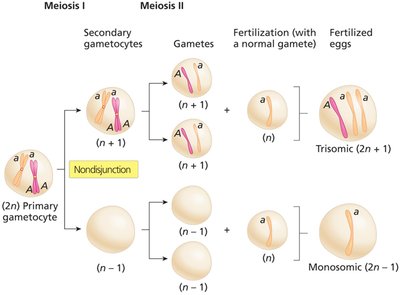
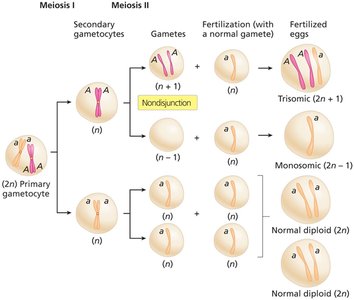
Nondisjunction and Aneuploidy
Nondisjunction is the failure of chromosomes or sister chromatids to separate properly during cell division, leading to cells with abnormal chromosome numbers (aneuploidy). This can occur during meiosis I or II, resulting in gametes with extra or missing chromosomes.
Euploid: Cells with complete sets of chromosomes (e.g., n, 2n, 3n).
Aneuploid: Cells with chromosome numbers not in complete sets (e.g., 2n+1, 2n-1).
Gene Dosage and Phenotypic Effects
Aneuploidy alters the dosage of all genes on the affected chromosome, often resulting in severe phenotypic consequences due to gene imbalance. Animals are generally more sensitive to gene dosage changes than plants.
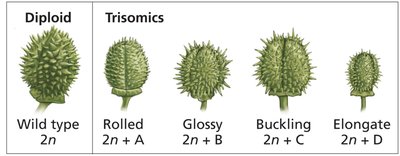
Aneuploidy in Humans
Most human aneuploidies are lethal, but some trisomies and sex chromosome aneuploidies are viable. The most common autosomal trisomies observed in live births are trisomy 13 (Patau syndrome), trisomy 18 (Edwards syndrome), and trisomy 21 (Down syndrome).
Aneuploidy | Syndrome | Frequency at Birth | Syndrome Characteristics |
|---|---|---|---|
Trisomy 13 | Patau syndrome | 1 in 15,000 | Mental retardation, developmental delay, major organ abnormalities, early death |
Trisomy 18 | Edwards syndrome | 1 in 6,000 | Mental retardation, developmental delay, skull and facial abnormalities, early death |
Trisomy 21 | Down syndrome | 1 in 1,500 | Mental retardation, developmental delay, characteristic facial abnormalities, short stature, variable life span |
Aneuploidy | Syndrome | Frequency at Birth | Syndrome Characteristics |
|---|---|---|---|
47, XXY | Klinefelter syndrome (males) | 1 in 1,000 | Variable secondary sexual characteristics, infertility, frequent breast swelling, no impact on mental capacity |
47, XYY | Jacobs syndrome (males) | 1 in 1,000 | Tall stature, common; possible reduction but not loss of fertility; no impact on mental capacity |
47, XXX | Triple X syndrome (females) | 1 in 1,000 | Tall stature, common; possible reduction of fertility, menstrual irregularity, no impact on mental capacity |
45, XO | Turner syndrome (females) | 1 in 5,000 | No secondary sexual characteristics; short stature; webbed neck common; no impact on mental capacity |
Down Syndrome and Maternal Age
The risk of Down syndrome (trisomy 21) increases with maternal age. This is likely due to the increased likelihood of meiotic nondisjunction in older oocytes.
Maternal Age Range | Total Live Births Studied | Trisomy 21 Births | Rate per 1000 Births |
|---|---|---|---|
15–19 | 30,272 | 18 | 0.49 |
20–24 | 117,593 | 87 | 0.73 |
25–29 | 108,746 | 96 | 0.90 |
30–34 | 49,487 | 72 | 1.56 |
35–39 | 19,522 | 72 | 3.69 |
40–44 | 4,880 | 73 | 18.02 |
45–49 | 304 | 19 | 55.02 |

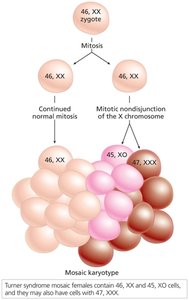
Mosaicism and Uniparental Disomy
Mosaicism results from mitotic nondisjunction early in embryogenesis, producing individuals with two or more genetically distinct cell lines. Uniparental disomy occurs when both copies of a chromosome are inherited from one parent, often due to trisomy rescue.
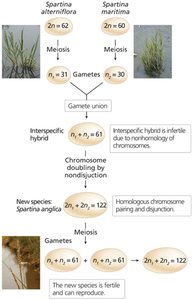
Polyploidy
Polyploidy is the presence of three or more complete sets of chromosomes. It can arise through autopolyploidy (chromosome duplication within a species) or allopolyploidy (combining chromosome sets from different species). Polyploidy is common in plants and can result in increased fruit size and hybrid vigor.
Odd-numbered polyploids (e.g., 3n, 5n) are often sterile, which is useful for producing seedless fruits.


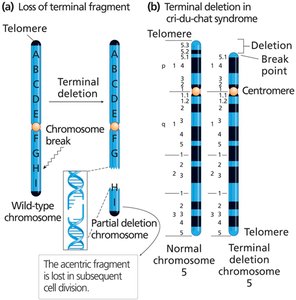
Chromosome Structural Mutations
Deletions and Duplications
Chromosome breakage can result in the loss (deletion) or gain (duplication) of chromosome segments. These mutations can have severe phenotypic effects due to gene dosage imbalances.
Terminal deletion: Loss of a chromosome end.
Interstitial deletion: Loss of an internal segment.
Partial deletion heterozygote: One normal and one deleted chromosome.
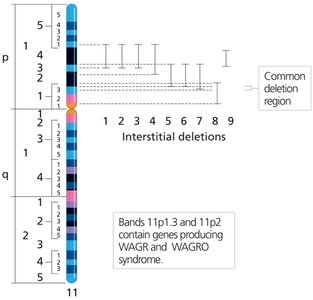
Unequal Crossover
Unequal crossing over during meiosis can produce one chromosome with a duplication and the other with a deletion. This is often associated with repetitive DNA sequences and can lead to genetic disorders such as Williams-Beuren syndrome.
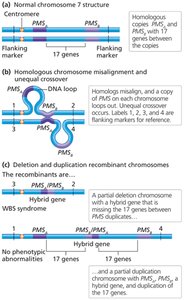
Detection of Deletions and Duplications
Large deletions or duplications can be detected by altered chromosome banding patterns. Microdeletions and microduplications require molecular techniques such as FISH for detection.
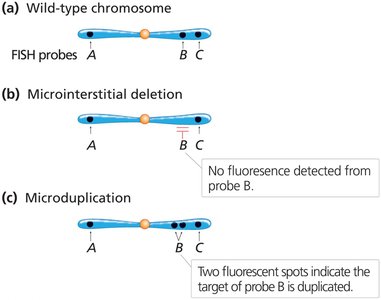
Microscopic Observation of Chromosome Loops
During prophase I of meiosis, large deletions or duplications can create unpaired loops when homologous chromosomes synapse. These loops are visible under the microscope and indicate regions of structural variation.
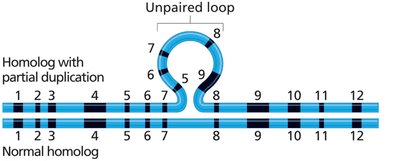
Deletion Mapping
Deletion mapping uses the phenomenon of pseudodominance to locate genes. If a dominant allele is deleted, a recessive allele on the homolog is expressed, revealing the gene's location.
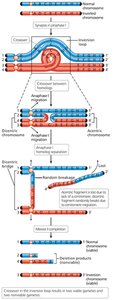
Inversions and Translocations
Chromosome breakage and incorrect reattachment can lead to inversions (reversal of a segment within the same chromosome) or translocations (exchange of segments between nonhomologous chromosomes).
Paracentric inversion: Centromere is outside the inverted region.
Pericentric inversion: Centromere is within the inverted region.
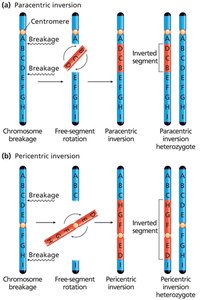
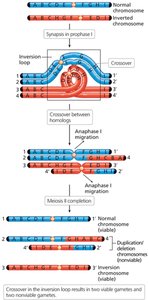
Consequences of Crossing Over in Inversion Heterozygotes
Crossing over within an inversion loop can produce chromosomes with duplications and deletions, leading to nonviable gametes. Paracentric inversions can produce dicentric (two centromeres) and acentric (no centromere) chromosomes, while pericentric inversions result in recombinant chromosomes with both duplications and deletions.
Translocations
Translocations involve the exchange of chromosome segments between nonhomologous chromosomes. There are three main types:
Nonreciprocal translocation: A segment moves to a nonhomolog without reciprocal exchange.
Reciprocal balanced translocation: Segments are exchanged between two nonhomologs.
Robertsonian translocation: Fusion of two acrocentric chromosomes, reducing chromosome number.
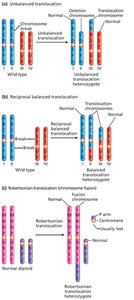
Segregation of Translocated Chromosomes
In reciprocal balanced translocation heterozygotes, chromosomes form a cross-shaped structure at metaphase I. Alternate segregation produces viable gametes, while adjacent segregation leads to unbalanced gametes and semisterility.
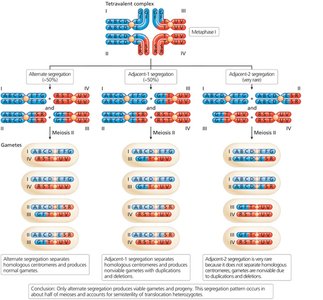
Robertsonian Translocation and Chromosome Evolution
Robertsonian translocations can reduce chromosome number and are a mechanism of karyotype evolution in related species.
Chromatin Structure and Higher-Order Organization
Chromatin and Histone Proteins
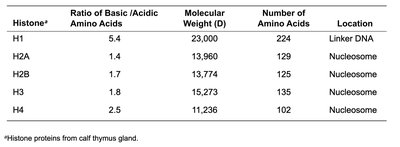
Eukaryotic chromosomes are composed of DNA and proteins, with histones being the major protein component. Histones organize DNA into nucleosomes, the fundamental units of chromatin structure.
Histone | Ratio of Basic/Acidic Amino Acids | Molecular Weight (D) | Number of Amino Acids | Location |
|---|---|---|---|---|
H1 | 5.4 | 23,000 | 224 | Linker DNA |
H2A | 1.4 | 13,960 | 129 | Nucleosome |
H2B | 1.7 | 13,774 | 125 | Nucleosome |
H3 | 1.8 | 15,273 | 135 | Nucleosome |
H4 | 2.5 | 11,236 | 102 | Nucleosome |
Nucleosome core particles consist of an octamer of histones (two each of H2A, H2B, H3, and H4) with ~146 bp of DNA wrapped around them. This structure compacts DNA about sevenfold.
Higher-Order Chromatin Structure
Chromatin fibers are further organized into higher-order structures:
10-nm fiber: "Beads-on-a-string" appearance (nucleosomes).
30-nm fiber: Solenoid structure stabilized by H1, with 6–8 nucleosomes per turn.
300-nm fiber: Loops of 30-nm fiber attached to a protein scaffold.
Chromatin compaction is essential for chromosome segregation during cell division and for regulating gene expression.
Position Effect Variegation (PEV)
PEV demonstrates that gene expression can be influenced by chromatin state. In Drosophila, moving a gene near heterochromatin can silence its expression in some cells, leading to variegated phenotypes.
Conclusion: Chromosome structure, number, and organization are central to genetics, affecting gene expression, inheritance, and evolution. Understanding these principles is essential for advanced study in genetics and genomics.
