 Back
BackGenetic Mechanisms: Double-Strand Break Repair, Transposable Elements, and Regulation of Gene Expression
Study Guide - Smart Notes
 Tailored notes based on your materials, expanded with key definitions, examples, and context.
Tailored notes based on your materials, expanded with key definitions, examples, and context.
Double-Strand Break Repair
Overview of Double-Strand Break Repair
Double-strand breaks (DSBs) in DNA are among the most dangerous types of genetic damage, potentially leading to chromosomal rearrangements, cancer, or cell death. Cells have evolved two primary pathways to repair DSBs: homologous recombination repair and nonhomologous end joining.
Homologous recombination repair: Uses a homologous sequence as a template for accurate repair.
Nonhomologous end joining: Directly ligates the broken DNA ends, often resulting in mutations.
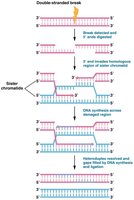
Homologous Recombination Repair
Homologous recombination repair is a high-fidelity mechanism that utilizes a sister chromatid as a template. It is most active during late S or early G2 phase when sister chromatids are available.
Recognition and processing: The break is recognized, and the 5' ends are digested, leaving 3' overhangs.
Strand invasion: The 3' overhang invades the homologous region of the sister chromatid.
DNA synthesis: DNA polymerase synthesizes new DNA using the sister chromatid as a template.
Resolution: The newly synthesized DNA is ligated, restoring the integrity of the chromosome.
Nonhomologous End Joining (NHEJ)
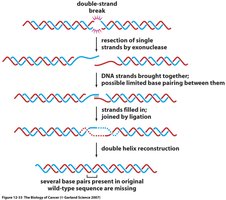
Nonhomologous end joining is a more error-prone repair mechanism, active primarily in G1 phase before DNA replication.
Protein complex formation: Proteins, including kinases and BRCA1, bind to the free DNA ends.
End processing and ligation: The ends are processed and directly ligated, often resulting in loss or addition of nucleotides.
Consequences: NHEJ can lead to mutations and chromosomal rearrangements.
Transposable Elements
Introduction to Transposable Elements
Transposable elements (TEs), also known as "jumping genes," are DNA sequences that can move within and between chromosomes. They are found in all organisms and contribute to genetic diversity and evolution.
Insertion sequences (IS elements): Simple transposons that can cause mutations by inserting into genes.
Bacterial transposons: Larger elements that can carry genes, such as those conferring drug resistance.
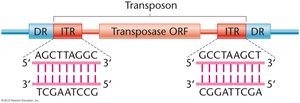
Structure of Transposons
Transposons typically contain inverted terminal repeats (ITRs) and direct repeats (DRs) flanking a transposase gene.
ITRs: Inverted sequences at the ends of the transposon.
DRs: Direct repeats generated during insertion.
Transposase: Enzyme required for movement.
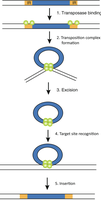
Mechanism of Transposition
Transposition involves several steps, including binding of transposase, excision, target site recognition, and insertion.
Transposase binding: Recognizes and binds to ITRs.
Complex formation and excision: Transposase forms a complex and excises the transposon.
Target site recognition: Transposase identifies a new insertion site.
Insertion: Transposon is inserted into the new site.
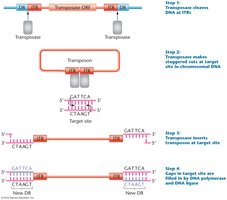
Transposon Insertion and Target Site Duplication
Transposase cleaves DNA at ITRs, makes staggered cuts at the target site, inserts the transposon, and fills gaps, creating new DRs.
Staggered cuts: Allow for insertion and duplication of target site sequences.
Gap filling: DNA polymerase and ligase fill gaps, completing the insertion.
Ac-Ds System in Maize
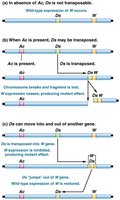
The Ac-Ds system in maize demonstrates the interaction between autonomous (Ac) and nonautonomous (Ds) elements.
Ac (Activator): Autonomous element that produces transposase.
Ds (Dissociation): Nonautonomous element requiring Ac for movement.
Phenotypic effects: Movement of Ds can disrupt the C gene, affecting kernel color.
Retrotransposons
Types of Retrotransposons
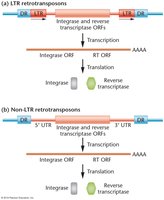
Retrotransposons are TEs that move via an RNA intermediate. They are classified as LTR (long terminal repeat) and non-LTR retrotransposons.
LTR retrotransposons: Contain long terminal repeats and encode integrase and reverse transcriptase.
Non-LTR retrotransposons: Lack LTRs but also encode integrase and reverse transcriptase.
Mechanism of Retrotransposition
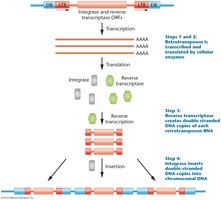
Retrotransposons are transcribed into RNA, translated to produce reverse transcriptase and integrase, and then inserted as DNA into the genome.
Transcription: RNA polymerase transcribes the retrotransposon.
Translation: Ribosomes produce reverse transcriptase and integrase.
Reverse transcription: Reverse transcriptase converts RNA to DNA.
Insertion: Integrase inserts the DNA into a new genomic location.
Transposable Elements in Drosophila
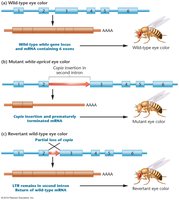
Copia Elements
Copia elements are a class of retrotransposons in Drosophila, characterized by direct terminal repeats (DTRs) and inverted terminal repeats (ITRs).
DTR: Direct terminal repeat, 267 bp long.
ITR: Inverted terminal repeat, 17 bp long.

Effects of Copia Insertion
Insertion of copia elements can disrupt gene function, such as altering eye color in Drosophila.
Wild-type: Normal gene and phenotype.
Mutant: Copia insertion disrupts gene, causing mutant phenotype.
Revertant: Partial loss of copia restores wild-type phenotype.
Regulation of Gene Expression in Eukaryotes
Levels of Regulation and Chromatin Remodeling
Gene expression in eukaryotes is regulated at multiple levels, including chromatin structure, transcription, RNA processing, and translation. Chromatin remodeling is essential for making DNA accessible to transcription machinery.
Chromatin structure: DNA is packaged with histones into nucleosomes.
Remodeling: Nucleosomes are repositioned or removed to allow access.
Chromatin Modifications
Eukaryotic DNA is combined with histones to form chromatin. Compact chromatin inhibits transcription, replication, and repair. Chromatin structure is dynamic and can be modified to regulate gene expression.
Histone modification: Covalent addition of acetyl, methyl, or phosphate groups to histone tails.
Acetylation: Decreases histone-DNA affinity, increasing transcription.
Methylation: Often suppresses transcription.
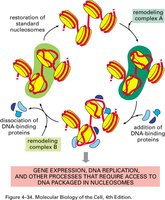
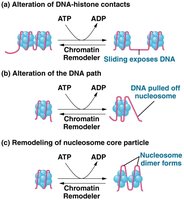
Chromatin Remodeling Complexes
SWI/SNF is a well-studied chromatin remodeling complex that loosens histone-DNA interactions and reorganizes nucleosome components.
Function: Makes DNA accessible to transcription factors and RNA polymerase II.
Gene Regulation: Promoters, Enhancers, and Silencers
Structure of Eukaryotic Genes
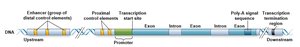
Eukaryotic genes contain coding regions, promoters, enhancers, silencers, and polyadenylation sites. Promoters act as the "on/off" switch for transcription.
Exons: Coding regions translated into protein.
Introns: Non-coding regions removed during RNA processing.
Promoter Structure and Diversity
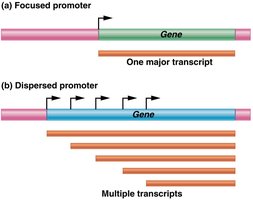
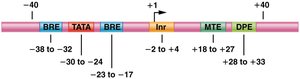
Promoters are made up of several DNA sequence elements, including the initiator (Inr), TATA box, BRE, DPE, and MTE. Promoters can be focused (single start site) or dispersed (multiple start sites).
Core promoter: Determines accurate initiation of transcription.
Proximal-promoter elements: Enhance basal transcription levels.
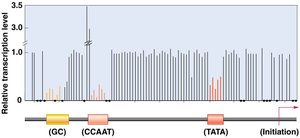
Proximal-Promoter Elements
Proximal-promoter elements, such as CAAT and GC boxes, are located upstream of the TATA box and enhance transcription.
Mutations: Can affect mRNA production levels.
Enhancers and Silencers
Enhancers and silencers are cis-acting elements that regulate transcription. Enhancers increase transcription, while silencers repress it.
Location: Can be upstream, downstream, or within the gene.
Function: Bind regulatory proteins to modulate gene expression.
Transcription Factors
Transcription factors are proteins that bind to cis-acting sites and regulate gene expression. They contain DNA-binding and trans-activating domains.
Activators: Increase transcription initiation.
Repressors: Decrease transcription initiation.
Common DNA Binding Motifs
Transcription factors often contain motifs such as zinc finger, helix-loop-helix, and leucine zipper, which facilitate DNA binding.
Zinc finger: Coordinates zinc ions for DNA binding.
Helix-loop-helix: Two α-helices connected by a loop.
Leucine zipper: Leucine residues form a zipper-like structure for dimerization.
Formation of RNA Polymerase II Initiation Complex
Pre-Initiation Complex (PIC)
General transcription factors assemble at the promoter to form the PIC, providing a platform for RNA polymerase II to recognize transcription start sites.
TFIID: Contains TATA binding protein (TBP) and TBP-associated factors (TAFs).
Additional factors: IIA, IIB, IIE, IIF, IIH.
Enhanceosome and Coactivators
Coactivators interact with activators and promoter-bound factors to form the enhanceosome, which facilitates transcription initiation.
Enhanceosome: Complex of proteins interacting with transcription machinery.
GAL Gene System in Yeast
Inducible Gene Expression
The GAL gene system in yeast is regulated by the presence of galactose. GAL genes are inducible and require activator proteins for transcription.
Structural genes: GAL1, GAL2, GAL7, GAL10 transport and metabolize galactose.
Regulatory genes: GAL3p, GAL4p, GAL80 regulate expression.
UASG: Upstream activating sequence acts as a regulatory element.
Posttranscriptional Regulation
Alternative Splicing
Alternative splicing generates different mRNA forms from identical pre-mRNA, increasing protein diversity. Introns are removed, and exons are spliced to produce mature mRNA.
Exons: Exit the nucleus and are translated.
Introns: Removed during splicing.
Spliceopathies: Disorders caused by mutations affecting splicing.
Example: Calcitonin Gene
The calcitonin gene produces two polypeptides via alternative splicing: calcitonin in the thyroid and CGRP in neurons.
Thyroid: Exons 1,2,3,4 produce calcitonin.
Neurons: Exons 1,2,3,5,6 produce CGRP.
Dscam Gene in Drosophila
The Dscam gene undergoes extensive alternative splicing, producing over 38,000 isoforms that guide axon growth and cell connections.
Isoforms: Unique combinations enable specific cell interactions.
Control of mRNA Stability
The steady-state level of mRNA is determined by transcription and degradation rates. mRNA stability is regulated by poly-A tail length, decapping, and endonuclease cleavage.
Poly-A tail: Stabilizes mRNA; shortening leads to degradation.
Decapping: Removal of 7-methylguanine cap destabilizes mRNA.
Endonuclease: Cleaves mRNA internally.
Protein Degradation: Ubiquitin and Proteasome
Proteins are tagged with ubiquitin for degradation by the proteasome, recycling amino acids.
Ubiquitin: Small protein that marks proteins for degradation.
Proteasome: Cylindrical structure that degrades tagged proteins.
p53 Protein Regulation
The p53 protein is a transcription factor that responds to DNA damage and stress. Its levels are tightly regulated by ubiquitin-mediated degradation.
Activation: DNA damage increases p53 levels.
Mdm2: Ubiquitin ligase that targets p53 for degradation.
Phosphorylation: Protects p53 from Mdm2, allowing it to activate transcription.
